Écriture : Thomas Buffet
Illustration : Pouaseuille
Relecture scientifique : Frédéric Chavane et Ali-Kémal Aydin
Relecture de forme : Geoffrey Pruvost et Eléonore Pérès
Temps de lecture : environ 20 minutes.
Thématiques : Neurobiologie (Biologie)
Publication originale : Berry M., et al., Anticipation of moving stimuli by the retina. Nature, 1999. DOI : 10.1038/18678
→ Des notions POUR APPROFONDIR à la fin de l’article

Le monde autour de nous est en mouvement permanent. Nos yeux transmettent des informations à notre cerveau, nous permettant de voir ce monde. Mais cette transmission se fait avec un certain délai. Nous devrions donc vivre dans le passé et en particulier voir les objets en mouvement à la position qu’ils occupaient un instant auparavant, en étant incapable de les attraper ou les éviter. Mais les auteurs de cet article scientifique ont montré que dans la rétine, la région de l’œil qui capte la lumière, un système particulier permet d’anticiper le mouvement des objets. Œil, cerveau, neurones… tout cela peut paraître compliqué. C’est pourquoi ce sera expliqué avec une histoire de reine et de châteaux.
Cet article décrit comment la rétine — c’est-à-dire le tissu nerveux situé au fond de l’œil qui transmet les informations lumineuses de notre environnement au cerveau sous forme d’impulsions électriques — peut anticiper le mouvement des objets. Mais avant de rentrer dans les détails de l’article, voici une histoire qui nous servira d’analogie pour mieux le comprendre.
Une histoire de reine et de châteaux
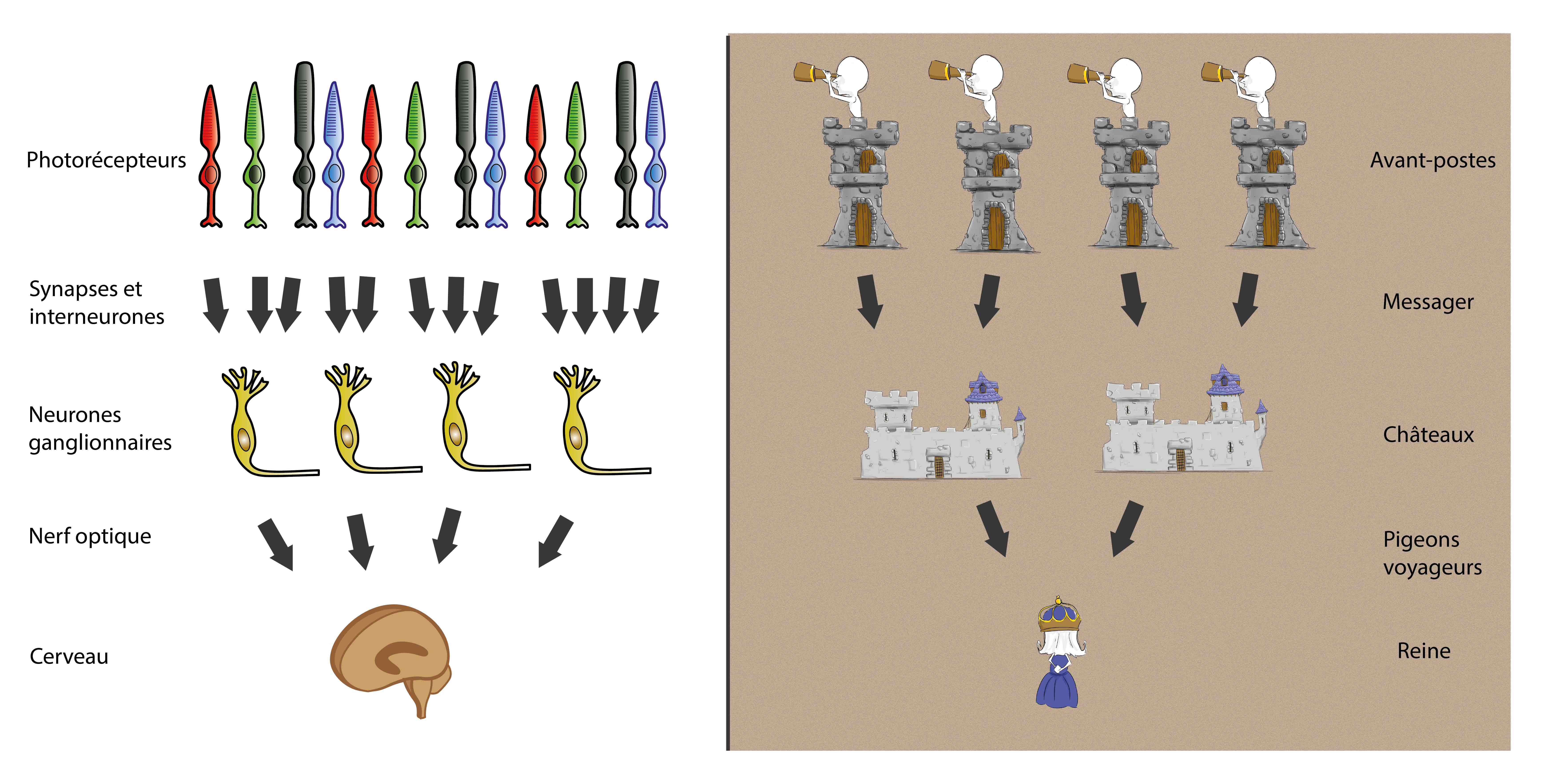
Imaginez un grand pays fantastique, où règne une reine aux longs cheveux blonds. Pour se protéger d’adversaires venus du Nord, elle a fait construire plusieurs châteaux à la frontière (Figure 1).
Afin de repérer au mieux les mouvements des armées ennemies, ces châteaux ont chacun des avant-postes dans la forêt, où des éclaireurs observent la frontière (Figure 2). Chaque jour, chaque avant-poste envoie un messager à son château pour porter un message simple : est-ce que l’avant-poste a aperçu ou non des ennemis. Ce trajet jusqu’au château prend au messager toute une journée et toute une nuit.

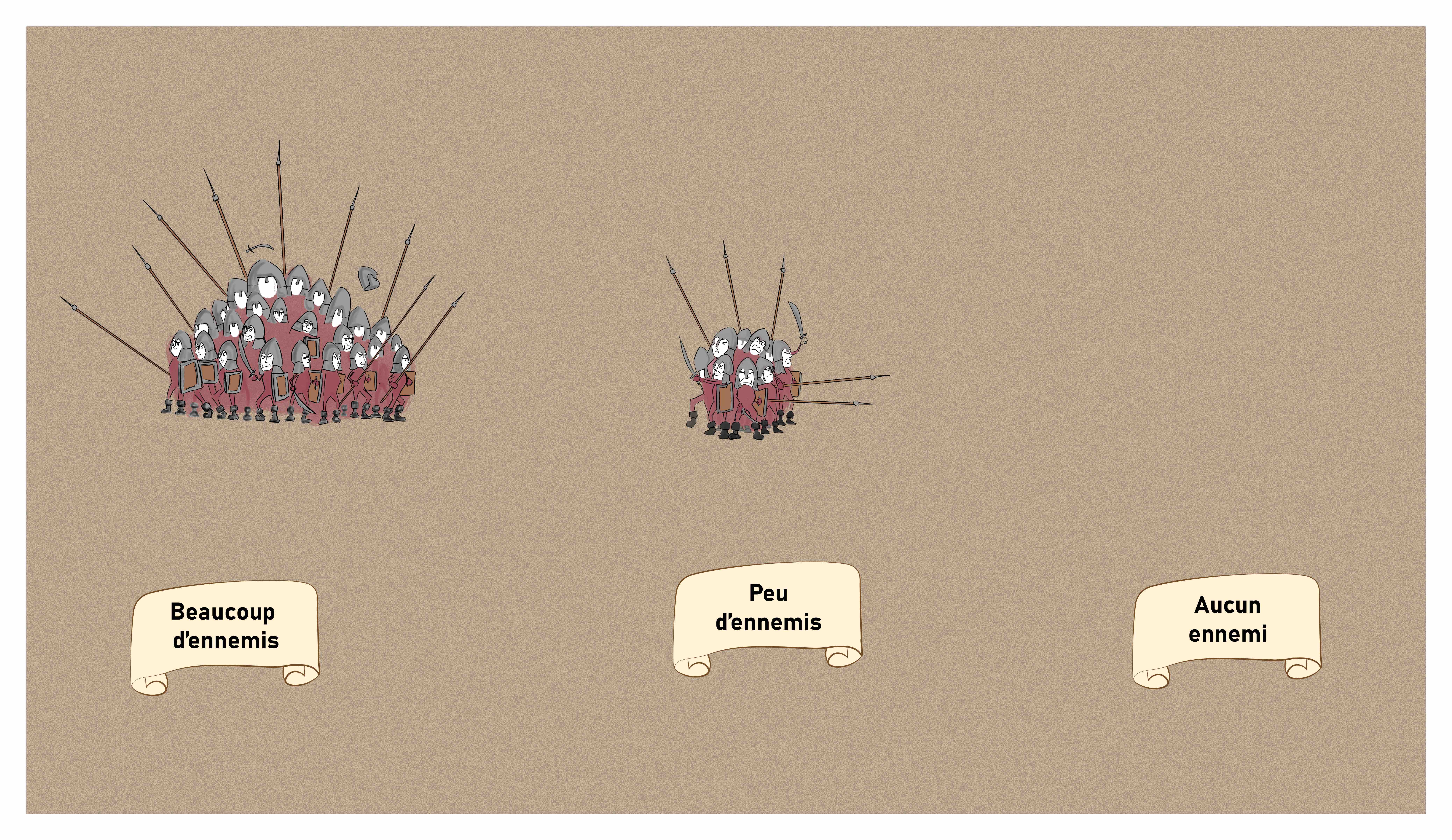
Une fois tous les messagers arrivés au château, le commandant en fait le bilan pour déterminer combien d’ennemis se trouvent en face de son château. Si aucun messager n’en rapporte, il n’y a pas d’ennemis. Si un seul en rapporte, il y a quelques ennemis et si tous les messagers en rapportent, il y a beaucoup d’ennemis (Figure 3).
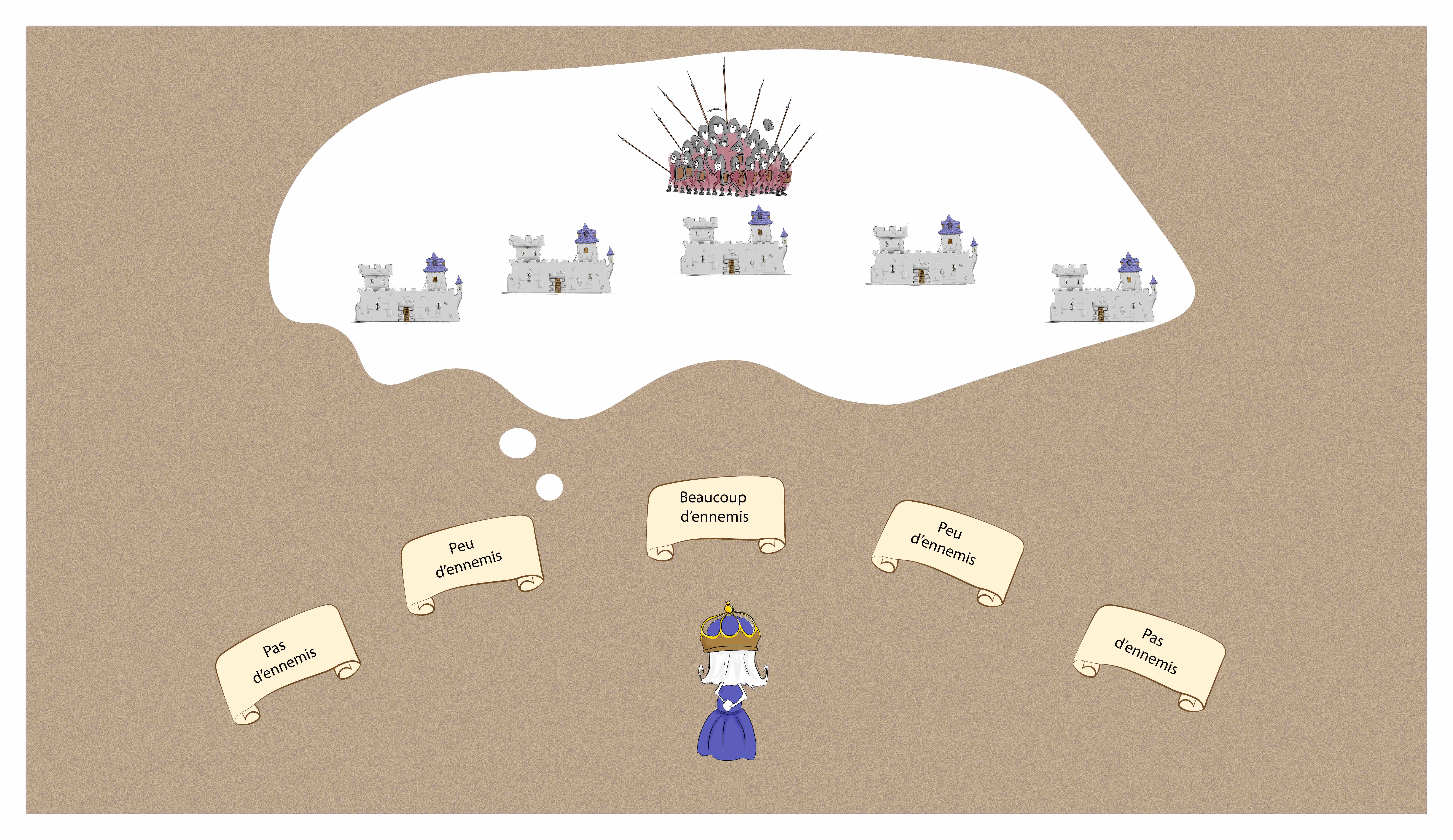
Le même manège se passe dans tous les châteaux et chaque commandant envoie ensuite un message à la reine par pigeon voyageur. Celui-ci fait le trajet en moins d’une heure. Lorsque la reine reçoit tous ces messages, elle peut connaître la taille et la position de l’armée ennemie (Figure 4).
La reine aimerait se débarrasser de ces armées ennemies qui rôdent derrière la forêt. Pour arriver à ses fins, elle possède un dragon qui peut voler jusqu’à la forêt en moins d’une heure et se débarrasser de l’armée. Un jour, elle reçoit des messages lui indiquant la position de l’armée. Elle y envoie son dragon. Mais surprise, il revient bredouille ! L’armée n’était plus là.
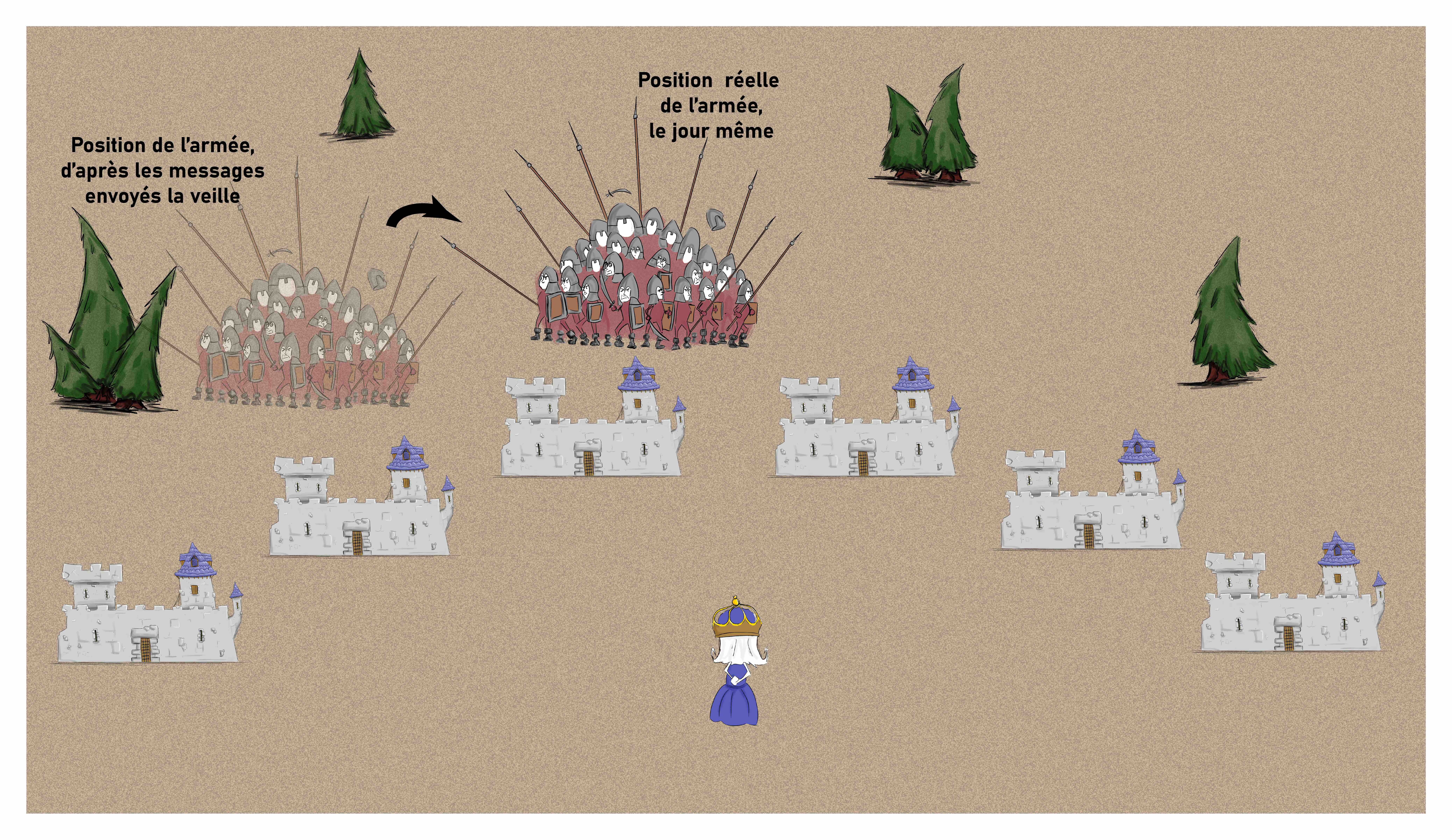
En effet, l’armée, qui se déplace d’ouest en est, parcourt en un jour la distance séparant deux châteaux. Mais le temps que la reine soit prévenue est plus long. Les messagers mettent en effet un jour et une nuit pour aller de leur avant-poste jusqu’au château. Il faut ensuite que les pigeons voyageurs atteignent la reine puis que le dragon vole dans l’autre sens. L’armée a largement eu le temps de se déplacer (Figure 5).
La reine ne se laisse pas décourager. La fois suivante, elle reçoit le premier jour des messages lui indiquant la présence d’ennemis. Elle n’envoie pas tout de suite le dragon mais attend le deuxième jour, puis le troisième. C’est seulement le troisième jour qu’elle l’envoie et le dragon trouve effectivement l’armée à la position qu’elle lui a indiquée.
Comment a-t-elle fait ? Laissons cette question en suspens le temps de nous intéresser à la publication scientifique.
L’anticipation du mouvement
Dans cet article, les scientifiques partent de deux constats. Le premier est que les informations que notre cerveau reçoit de nos organes sensoriels — comme les yeux — sont en retard sur la réalité. En effet, il faut un certain temps à ces organes sensoriels pour générer ces informations, puis pour les transmettre le long des différentes étapes du système visuel de notre cerveau. À un instant donné, le cerveau reçoit donc les informations qui ont été générées un instant plus tôt. Ce premier constat est généralement résumé par la phrase « nous vivons dans le passé ». Mais est-ce vraiment le cas ?
Le deuxième constat est que l’être humain est néanmoins capable d’interagir correctement avec son environnement, et en particulier avec des objets en mouvement, pour les attraper ou les éviter. Cette capacité repose sur la motricité, mais également sur la perception. En effet, nous sommes capables, dans une certaine mesure, d’anticiper la trajectoire des objets en mouvement pour placer notre corps à un endroit approprié. Mais dès le stade de la perception, une anticipation est présente. Cela a été montré dans des études précédentes [1], notamment grâce à l’illusion nommée effet flash-lag. Lorsqu’une barre est montrée très brièvement (flashée) alors qu’une autre barre se déplace en même temps, une perception étrange apparaît (Figure 6).

La barre en mouvement est perçue un peu à droite de la barre flashée. Pourtant, si l’on reprend le film image par image, la barre flashée apparaît exactement en face de la barre en mouvement.
En 1999, lorsque cet article scientifique a été publié, la communauté scientifique considérait plutôt que d’autres régions du cerveau étaient responsables de l’anticipation, et non les yeux, que ce soit la partie motrice ou la partie perceptive. Mais les auteurs remettent en cause cette explication. Ils affirment que cette anticipation se déroule au moins en partie déjà au niveau de l’œil et plus précisément de la rétine, la partie de l’œil qui transmet des informations aux autres parties du cerveau. Selon les auteurs, la rétine est plus qu’une simple caméra en retard sur la réalité. Avant d’expliquer comment ils ont prouvé cette capacité étonnante de la rétine, il est nécessaire de bien comprendre comment celle-ci fonctionne.
Le fonctionnement de la rétine
Les connaissances sur le système visuel nous apprennent que la lumière de notre environnement entre dans l’œil en traversant la cornée puis le cristallin, qui la font converger vers la rétine (Figure 7).
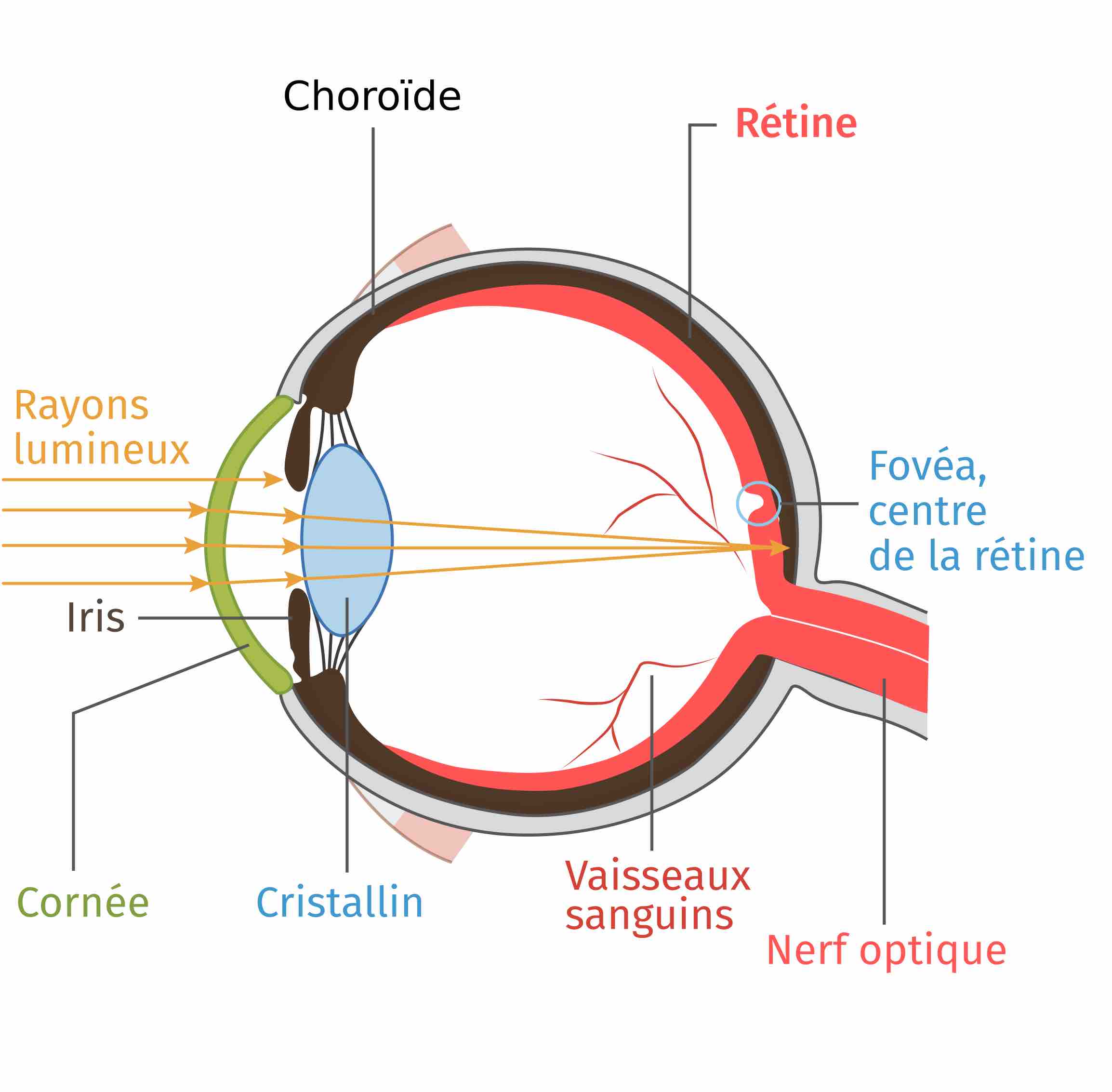
La rétine est un tissu nerveux constitué principalement de neurones, des unités qui s’échangent des informations au niveau de synapses, les points de rencontres entre plusieurs neurones. Des neurones particuliers, les photorécepteurs, commencent par transformer la lumière en signaux électriques. Puis ils transmettent ces signaux par plusieurs synapses jusqu’à d’autres neurones de la rétine, les cellules ganglionnaires. À ce niveau, les informations sont rassemblées et relayées par le nerf optique jusqu’au reste du cerveau, qui les interprète et peut coordonner des mouvements et un comportement adaptés (Figure 8). La rétine est en réalité composée d’autres types de neurones, mais ils seront ignorés dans ces explications pour plus de simplicité.
La rétine transmet vers le cerveau, entre autres, la position des objets dans le champ visuel. Pour déterminer ces positions, le cerveau rassemble les informations de plusieurs cellules ganglionnaires. Ce fonctionnement est similaire à celui de l’histoire. Les éclaireurs créent une information sur l’armée ennemie, qui est d’abord rassemblée par les châteaux, puis envoyée à la reine. Celle-ci détermine la position de l’armée en rassemblant les informations de plusieurs châteaux (Figure 8).
L’expérience
Comme expliqué précédemment, la position des objets en mouvement est perçue correctement malgré le temps que met l’information à parvenir au cerveau. Pour expliquer cette observation, au moins en partie, les auteurs ont une hypothèse. Ils pensent que l’information nécessaire pour anticiper le déplacement d’un objet en mouvement serait déjà présente au niveau des neurones ganglionnaires. Il y aurait ainsi une anticipation du mouvement directement au niveau de la rétine ! Pour la démontrer, ils ont réalisé une expérience sur la rétine du lapin et de la salamandre, deux modèles animaux [pour approfondir : voir note A].
Dans cette expérience, les auteurs utilisent une technique d’électrophysiologie : l’enregistrement de l’activité électrique des neurones. En effet, les informations transmises par les neurones sont sous forme d’impulsions électriques (les potentiels d’actions). Étudier cette activité électrique peut nous aider à comprendre le message que transmettent les neurones. À l’aide d’électrodes, ils enregistrent donc l’activité des neurones ganglionnaires de la rétine qui transmettent les informations électriques au cerveau par le nerf optique. En même temps, ils utilisent un micro-projecteur pour éclairer la rétine. De cette façon, les auteurs peuvent simuler le déplacement d’un objet devant l’œil tout en enregistrant l’activité neuronale générée par ce mouvement [pour approfondir : voir note B].
Dans notre histoire, cela reviendrait à simuler le mouvement d’une armée dans la forêt et à intercepter les pigeons voyageurs des châteaux pour comprendre quelles informations ils envoient à la reine.
Les auteurs utilisent leur micro-projecteur pour simuler deux types d’images. La première est une barre lumineuse flashée qui simule un objet apparaissant et disparaissant brusquement. La deuxième est une barre se déplaçant de gauche à droite ou inversement simulant un objet en mouvement. Ils peuvent ainsi recréer un cadre similaire à celui de l’illusion d’optique présentée plus haut (Figure 6). Ils analysent ensuite comment les neurones ganglionnaires réagissent à ces deux types d’images.
Réponse individuelle d’un neurone à un flash lumineux
La première étape est de vérifier que leurs modèles ont le même comportement que les données déjà existantes concernant le délai entre un flash lumineux et la réaction des neurones ganglionnaires. En enregistrant l’activité d’un seul neurone ganglionnaire, les auteurs confirment la notion de retard de l’information visuelle par rapport à la réalité. En effet, si le flash apparaît à un instant donné, ils observent que la réaction d’un neurone ganglionnaire survient environ 50 millisecondes (ms) plus tard (Figure 9).
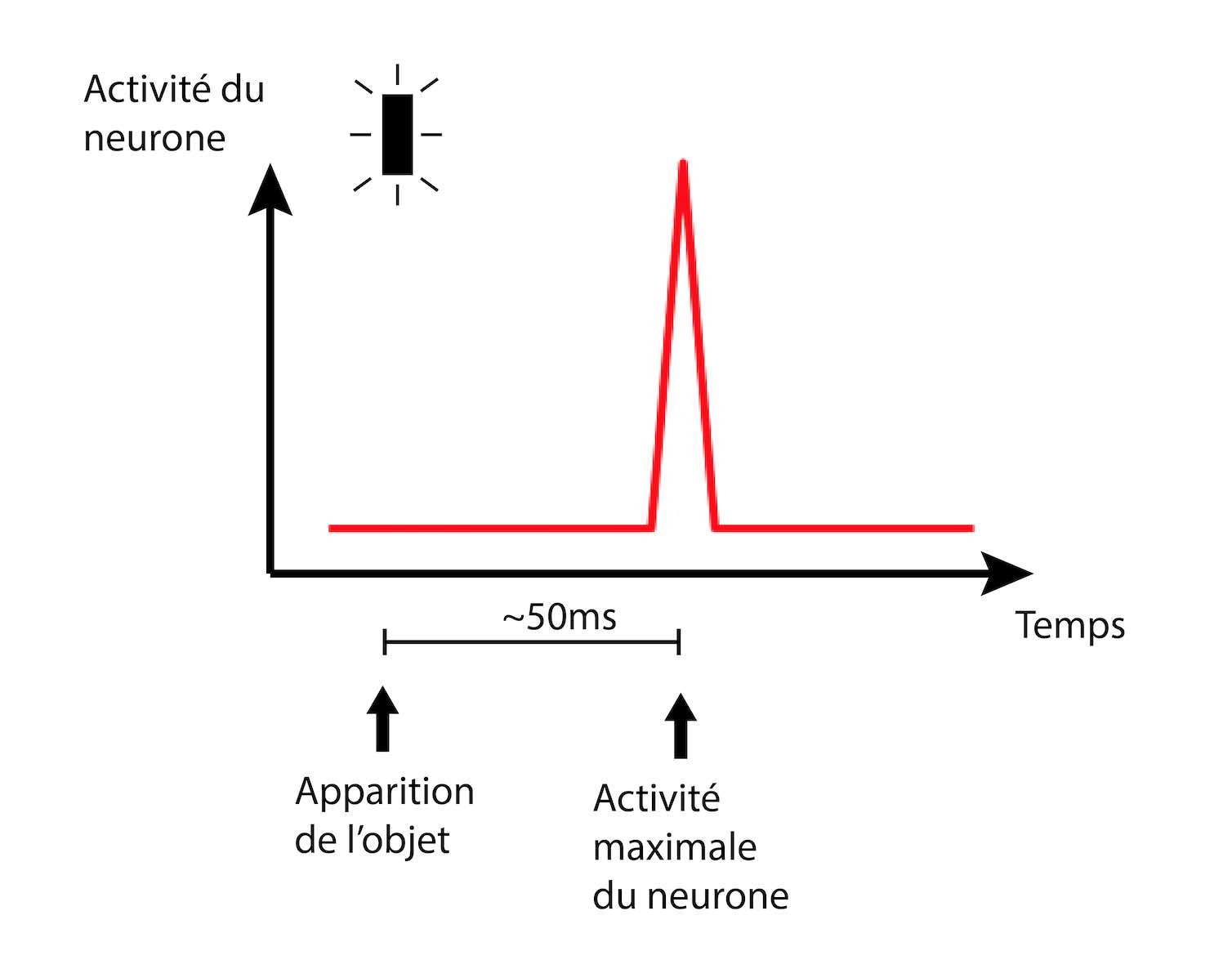
Dans notre histoire, cela reviendrait à constater qu’un château envoie un pigeon voyageur à la reine avec une journée de retard, à partir du moment où l’éclaireur a vu l’armée ennemie, à cause du temps de trajet nécessaire aux messagers.
Mais dans notre histoire, l’armée se déplace. Comment réagit un neurone à un objet en mouvement ?
Réponse d’un neurone à un objet en mouvement
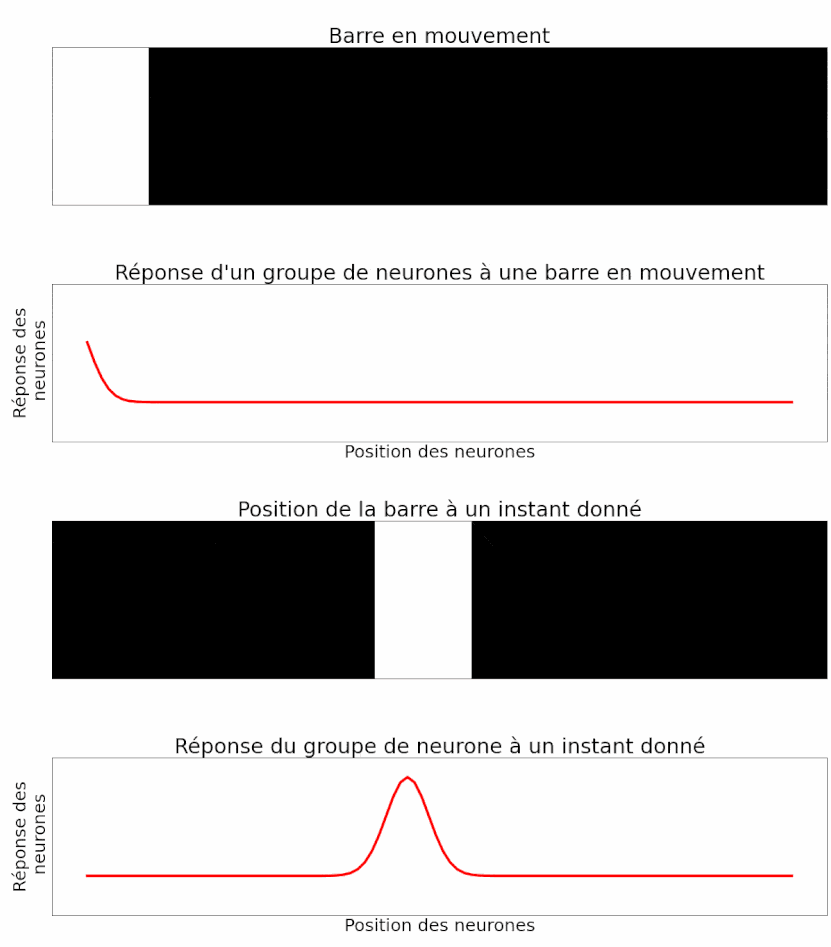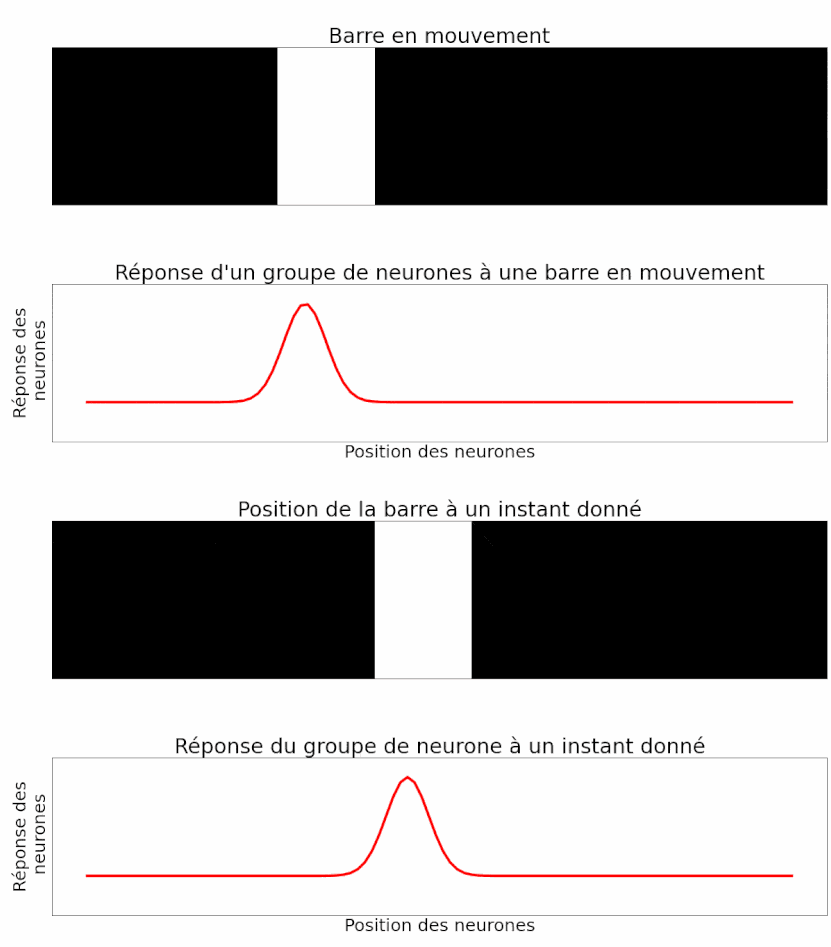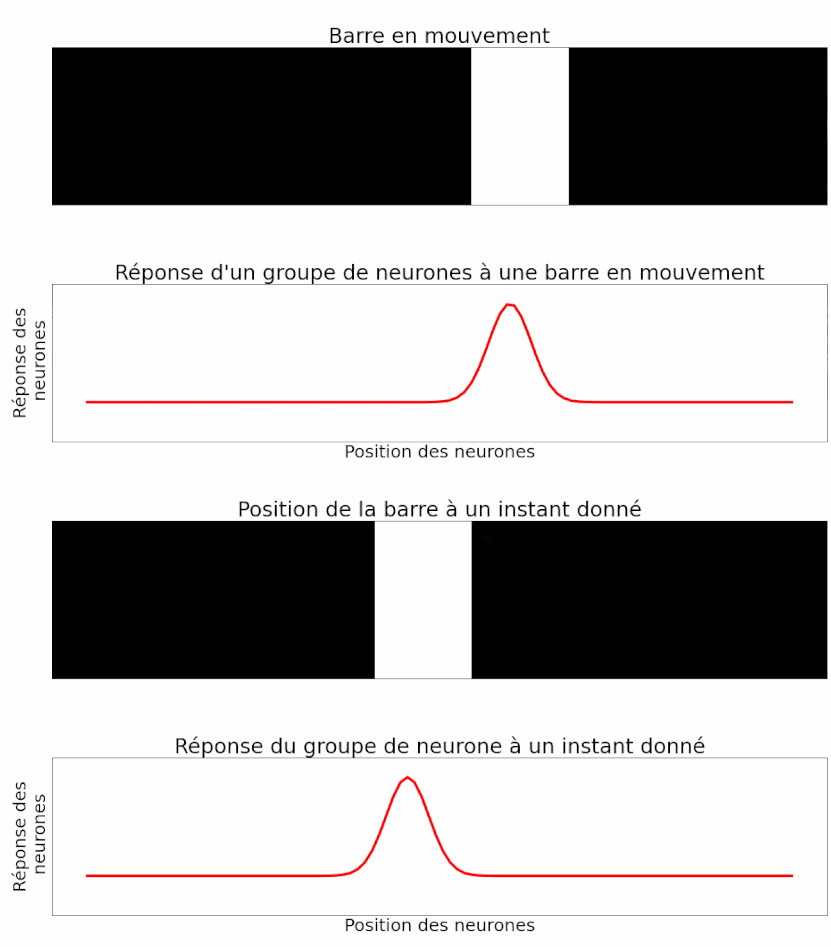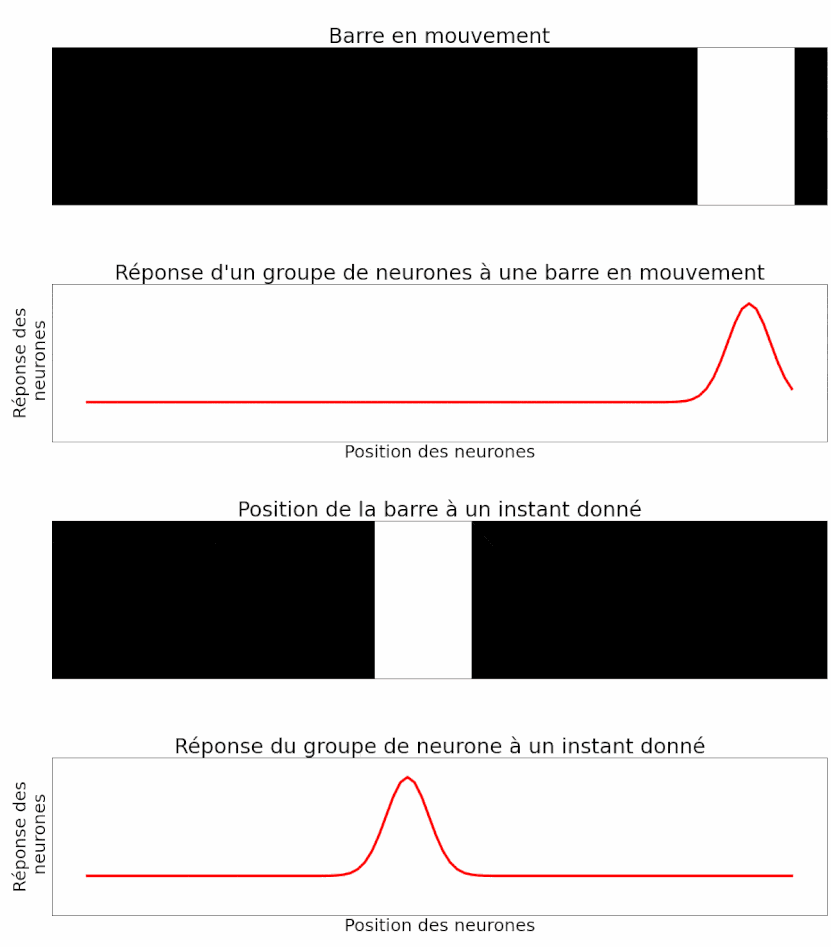
Lorsqu’un objet traverse le champ visuel, la réponse du neurone est un peu différente. Il commence par réagir faiblement lorsqu’une partie seulement de l’objet est à sa portée. Puis, il réagit au maximum lorsque l’objet est en face. À nouveau, il réagit plus faiblement lorsque l’objet s’éloigne avant de s’éteindre complètement (Figure 10) [pour approfondir : voir note C].
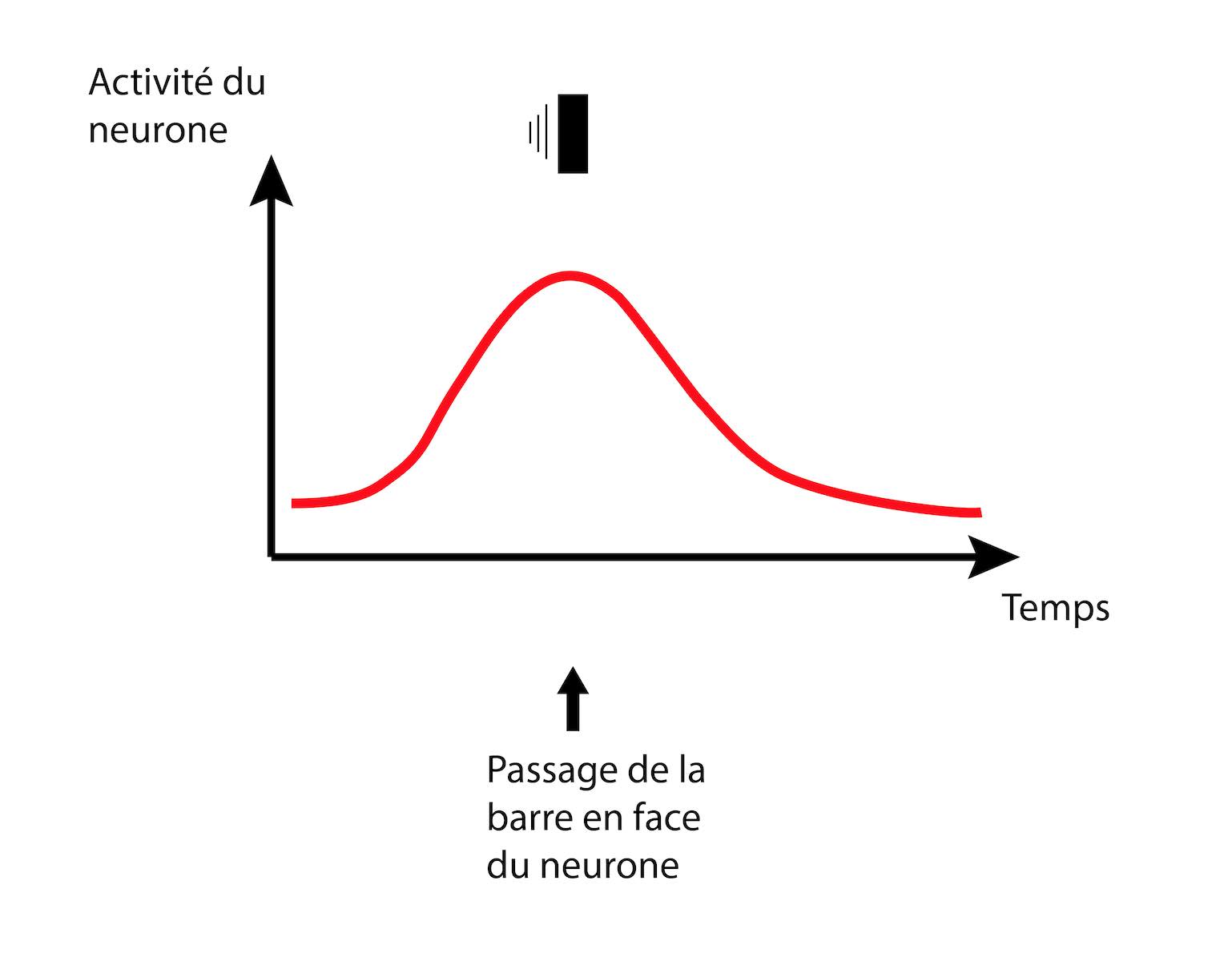
On constate que la réponse d’un neurone est bien différente dans le cas du mouvement. On observerait le même comportement pour un château dans notre histoire : l’activité augmente au fur et à mesure qu’une armée approche, puis décroît lorsqu’elle s’éloigne.
Mais dans l’histoire, la reine ne se contente pas d’un seul château. Elle en a plusieurs et combine les informations pour connaître la position de l’armée. Que se passe-t-il lorsque l’on s’intéresse à l’activité d’une population de neurones, au lieu d’un neurone isolé, pour ces deux mêmes types d’images ?
Réponse d’un groupe de neurones à un flash
La position d’un objet est déterminée par le cerveau à partir de l’activité d’un groupe de neurones ganglionnaires, et non d’un neurone isolé. Les auteurs analysent donc ensuite l’activité d’un groupe de neurones au même flash de lumière : ils retrouvent la même propriété de retard chez tous ces neurones. Ils montrent également une autre propriété de la réponse collective à un flash : les neurones situés exactement en face du flash répondent très fortement, alors que les neurones situés sur les côtés, plus loin, répondent moins fortement (Figure 11). En effet, le flash n’est pas entièrement dans leur champ récepteur, la partie du champ visuel à laquelle chaque neurone est sensible.
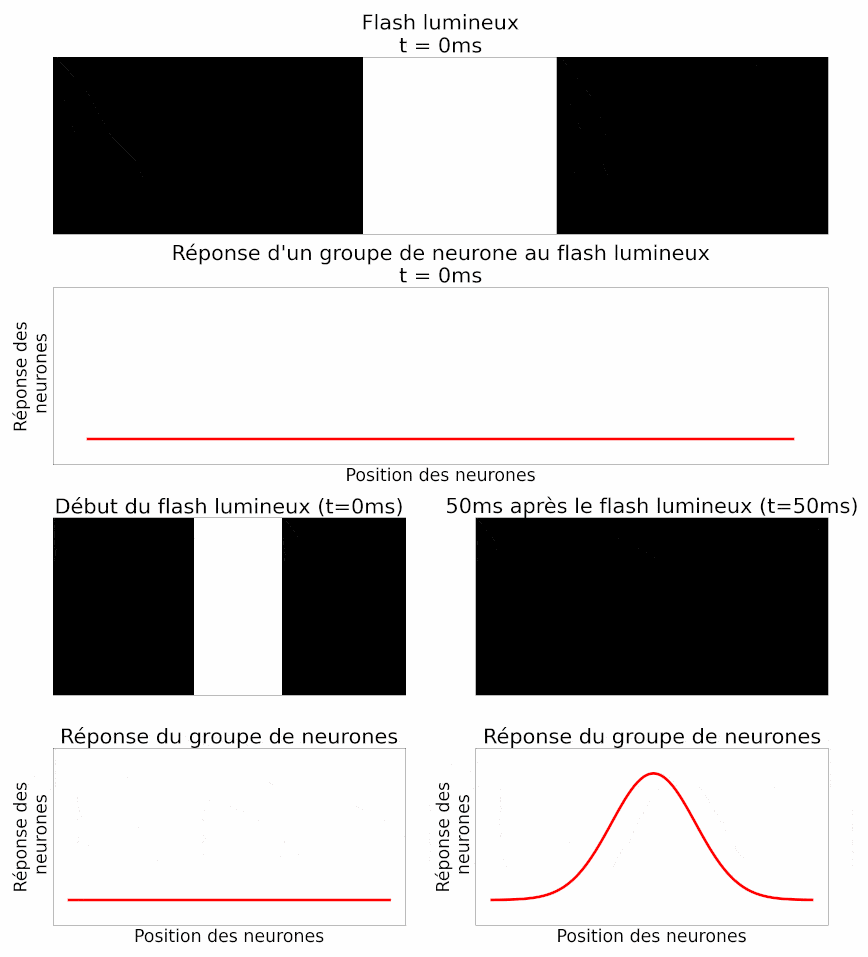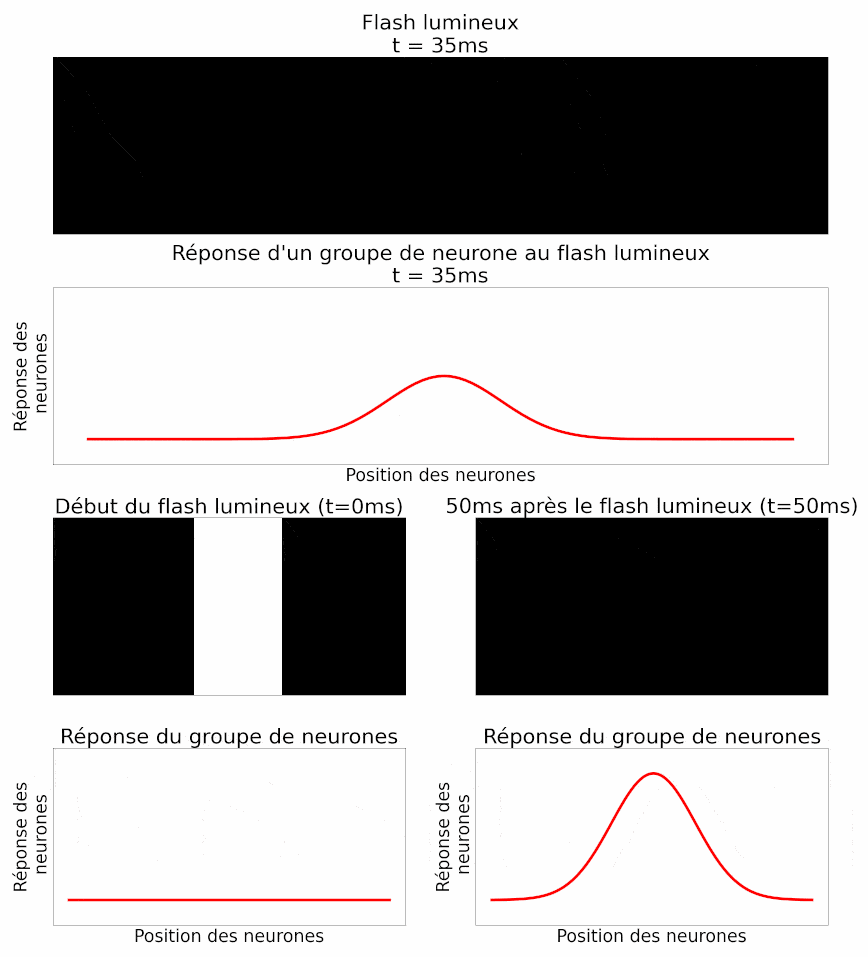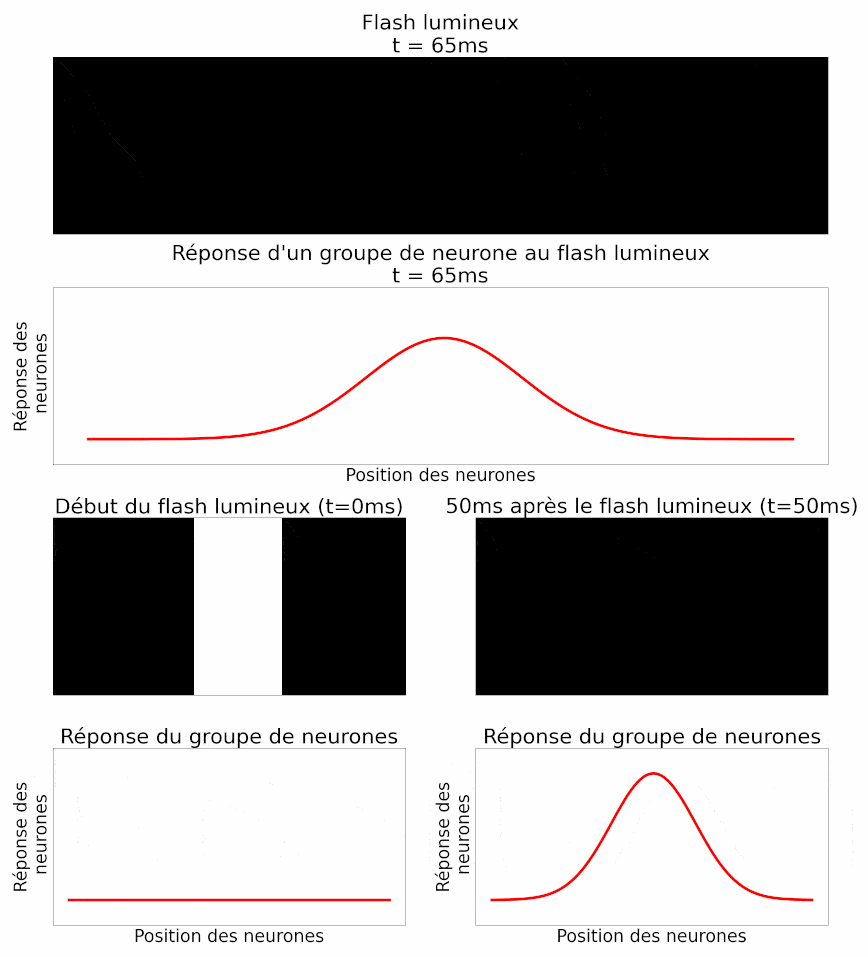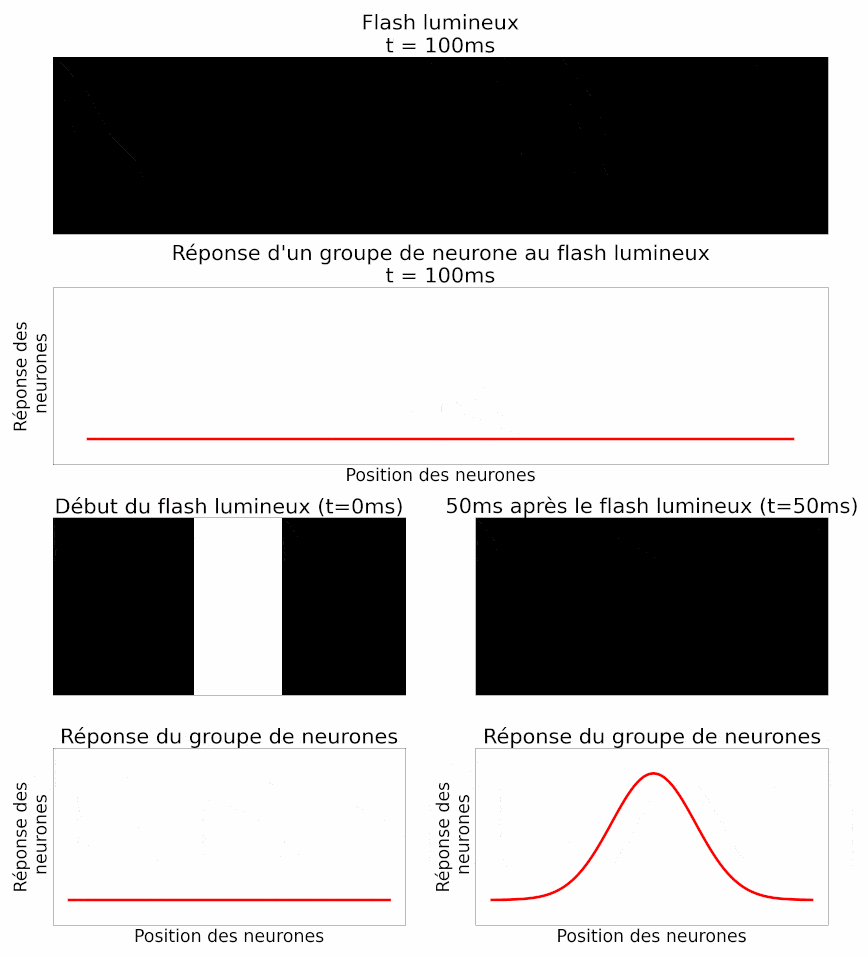
Dans l’histoire, cela équivaut à des châteaux plus éloignés de la position de l’armée, qui signaleraient moins d’ennemis car un plus petit nombre d’éclaireurs l’aurait vue.
Enfin, comment notre population de neurones réagit-elle à un objet en mouvement ?
Réponse d’un groupe de neurones à un objet en mouvement
Les auteurs peuvent mesurer la réponse d’un groupe de neurones à un objet en mouvement. Et c’est là que leur étude est la plus intéressante ! Étant donné ce qu’ils ont montré précédemment, les auteurs sont capables d’interpréter la réponse du groupe de neurones pour déterminer la position du flash. Ils peuvent procéder de la même manière, mais cette fois-ci avec un objet en mouvement. On sait que la réponse d’un groupe de neurones au flash est temporellement en retard par rapport à son apparition (Figure 11). La réponse à un objet en mouvement devrait donc faire de même : en signalant la position de la barre en mouvement avec environ 50 ms de retard. Mais le résultat est différent. En réalité, lorsque l’on analyse la réponse d’un groupe de neurones à un instant donné, celle-ci décrit effectivement la position de l’objet à cet instant. Il n’y a pas de retard (Figure 12) ! Il y a donc bien une anticipation de la trajectoire d’objets en mouvement par les cellules ganglionnaires de la rétine. Ou du moins une correction des délais de traitement et de transmission [pour approfondir : voir note D].
Pour mieux étudier ce phénomène et vérifier qu’il ne s’agit pas juste d’un cas particulier, plusieurs aspects de l’image utilisée ont été modifiés et testés [pour approfondir : voir note E].
Avec cette expérience, les auteurs ont donc démontré que l’activité groupée des cellules ganglionnaires permet de compenser les délais de transmission inhérents à la rétine pour transmettre une information correcte sur la position d’un objet en mouvement, le point de référence étant la réponse retardée à un flash.
Mais comment cela est-il possible ?
Le mécanisme derrière l’anticipation
Nous voilà donc arrivés à la même question, dans la publication scientifique et dans notre histoire. Comment est-il possible, à partir d’unités transmettant avec un certain retard des informations sur la position d’un objet, de retrouver la position exacte d’un objet à un instant donné ?
Commençons par notre reine et ses châteaux. Comment a-t-elle pu connaître la position exacte de l’armée ennemie pour y envoyer son dragon ?
Des messages modifiés
Voyons une explication possible. La reine a envoyé aux commandants de tous les châteaux de nouvelles consignes sur la façon dont ils devaient lui transmettre les informations par pigeon voyageur. Ils devaient non seulement rassembler les informations de leurs différents éclaireurs, mais aussi adapter le message en fonction de ce qu’ils avaient vu le jour précédent, avec une règle simple, pouvant être résumée par la Figure 13 suivante.
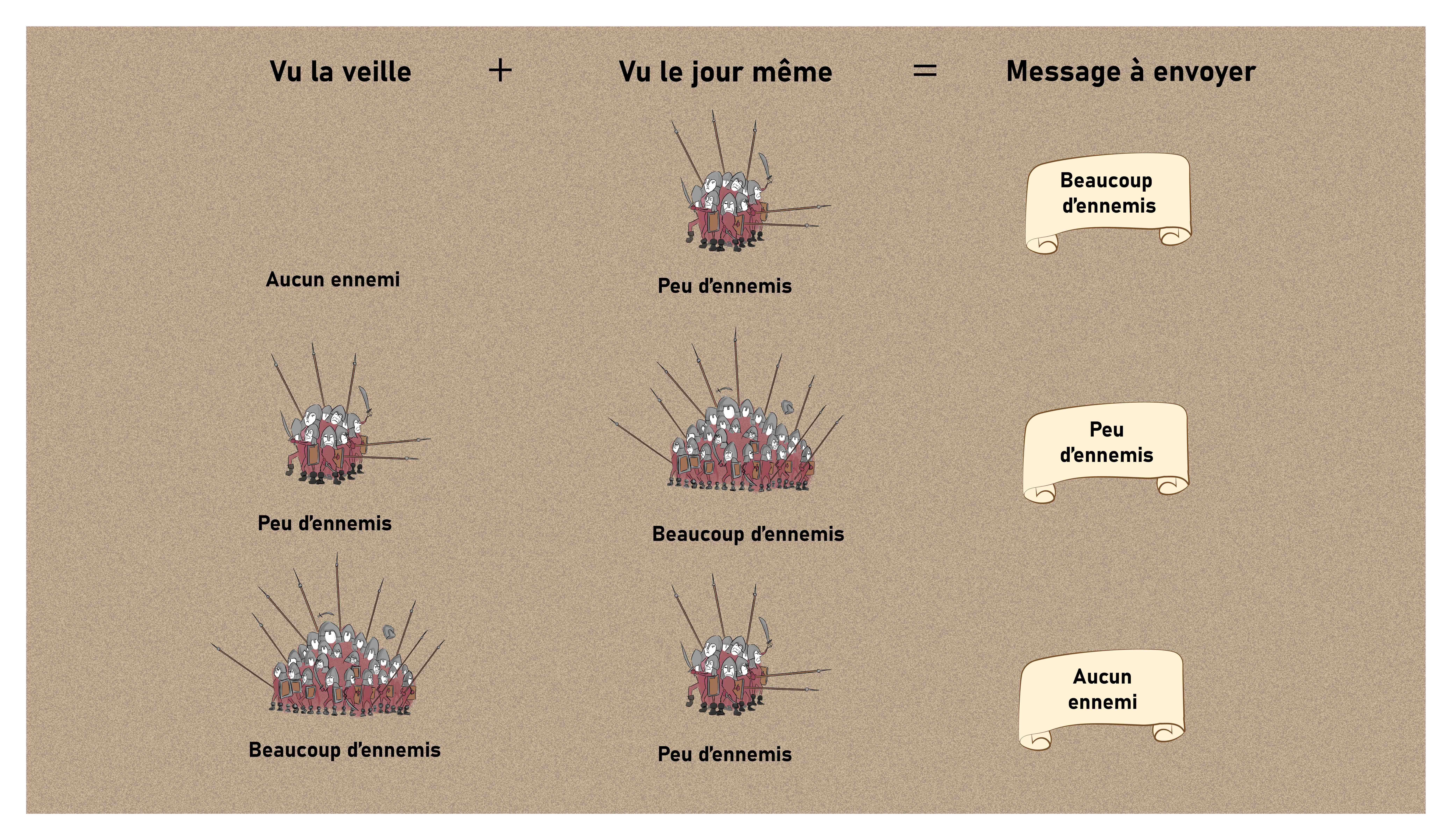
Arrêtons-nous un instant sur les modifications apportées par ces nouvelles consignes. À l’échelle d’un unique château, cela va avoir l’effet d’exagérer la quantité d’ennemis signalés lorsque ceux-ci arrivent à proximité (Figure 13, ligne 1). À l’inverse, lorsque les ennemis sont déjà visibles et deviennent simplement plus nombreux ou disparaissent, leur nombre est minimisé (Figure 13, lignes 2 et 3).
Cela devient intéressant lorsque l’on combine les informations de plusieurs châteaux pour retrouver la position de l’armée. Avec les messages originaux, l’armée est localisée devant le château devant lequel elle se trouve, mais cette information devient erronée le temps de la transmettre puisque l’armée se déplace (Figure 14).
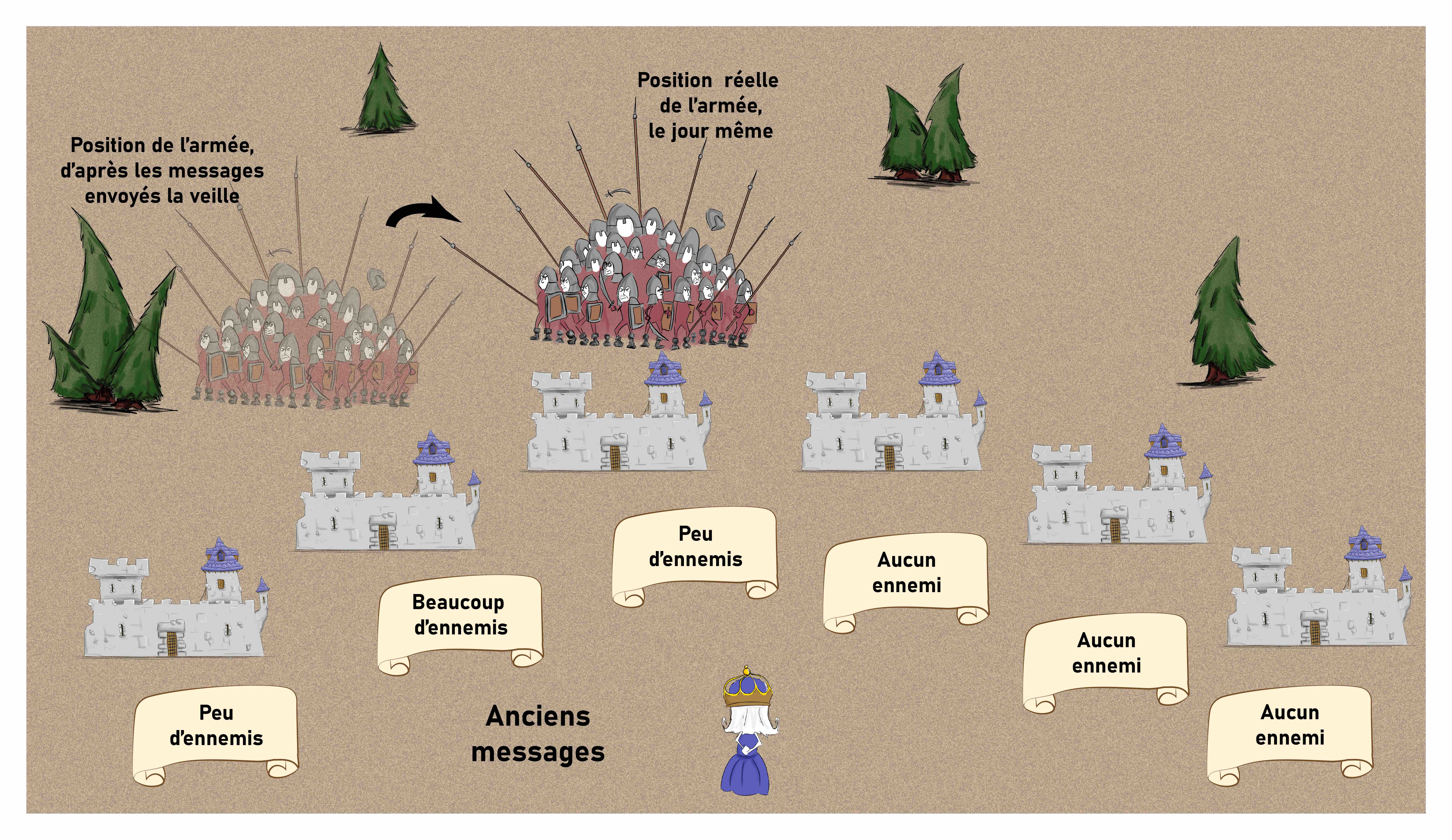
Avec les nouvelles consignes en revanche, l’armée est signalée très rapidement lorsqu’elle arrive en vue d’un nouveau château, alors que les châteaux qui la voient s’en aller commencent à l’ignorer. Ce système permet donc d’anticiper légèrement la position future de l’armée, et donc de contrebalancer les délais de transmission (Figure 15).
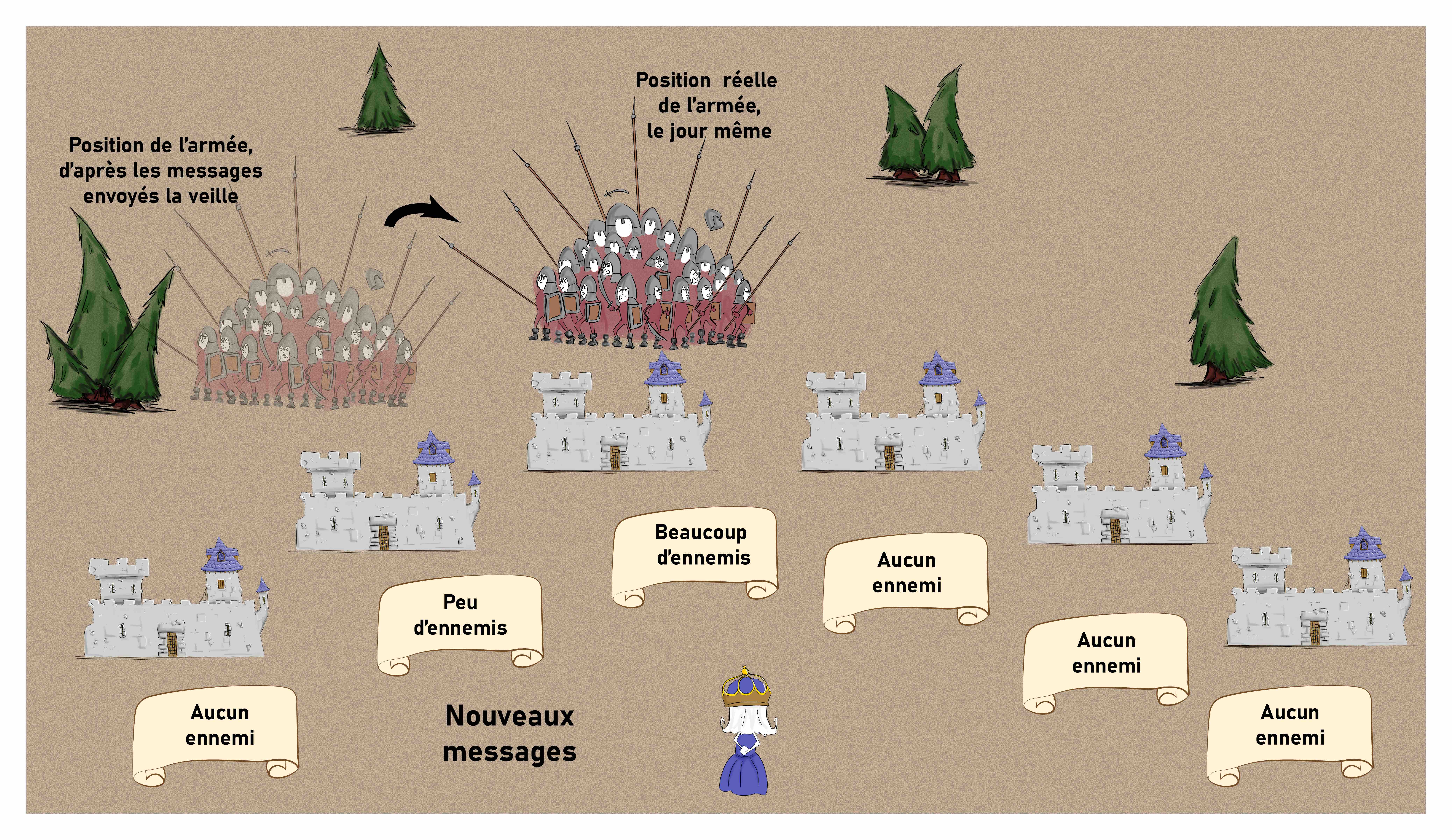
Il est important de noter que la reine ne modifie pas sa façon d’interpréter les messages. Ce sont les châteaux frontaliers qui envoient des messages qui ne sont pas exactement conformes à la réalité pour que la reine puisse prendre les bonnes décisions [pour approfondir : voir note F].
L’anticipation par les neurones
Un mécanisme similaire a été proposé pour les neurones ganglionnaires de la rétine, nommé contrôle de gain. Le principe général est le même que celui de la modification des messages par la reine. Lorsqu’un objet, comme une barre lumineuse en mouvement, commence à stimuler un neurone, celui-ci réagit très fortement. Mais alors que cette stimulation continue, voire s’amplifie, l’activation du neurone est réduite. Et lorsque la stimulation se termine, l’activation du neurone disparaît très rapidement. L’activité d’un neurone dépend donc à la fois de ce qui est présent devant lui à un instant donné, mais également du passé récent. Ce principe peut être résumé de la même façon dans un tableau (Figure 16).
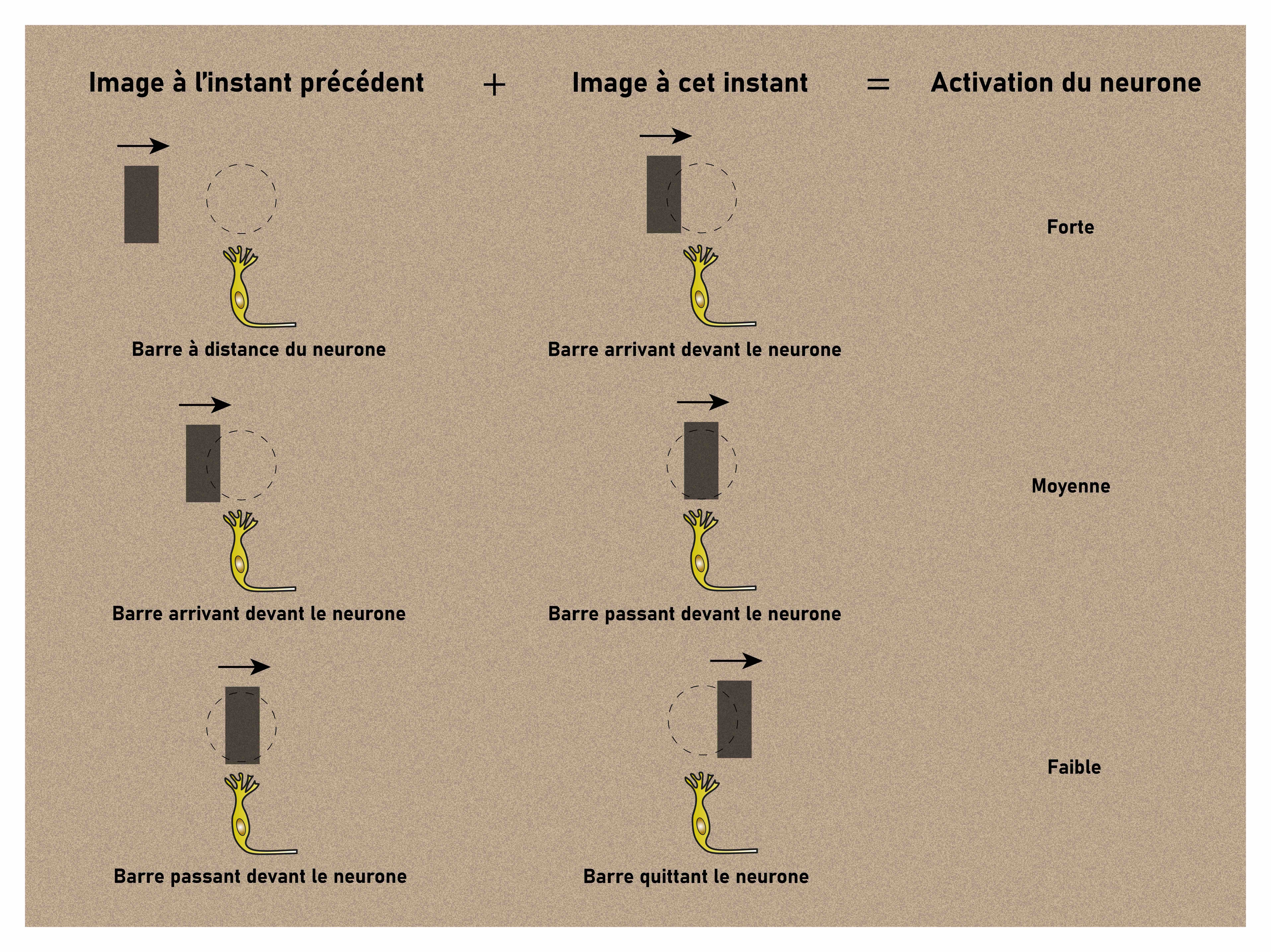
Pour tester cette hypothèse, les auteurs ont construit un modèle informatique représentant un groupe de neurones ganglionnaires possédant ou non cette capacité de contrôle de gain. Dans le modèle informatique, tout est beaucoup plus précis. En effet, on n’utilise pas des cas séparés mais des fonctions mathématiques qui déterminent, pour chaque succession de stimulation visuelle, quelle sera la réponse exacte du neurone.
À partir de ce modèle, les auteurs ont simulé la réponse des neurones ganglionnaires à différentes stimulations visuelles, comme une barre en mouvement. Leur observation est la suivante : si les neurones modélisés possèdent cette capacité de contrôle de gain, la position prédite de la barre en mouvement est correcte (Figure 17, en rouge), mais si on retire cette capacité aux neurones, la position prédite souffre du retard de transmission (Figure 17, en gris).
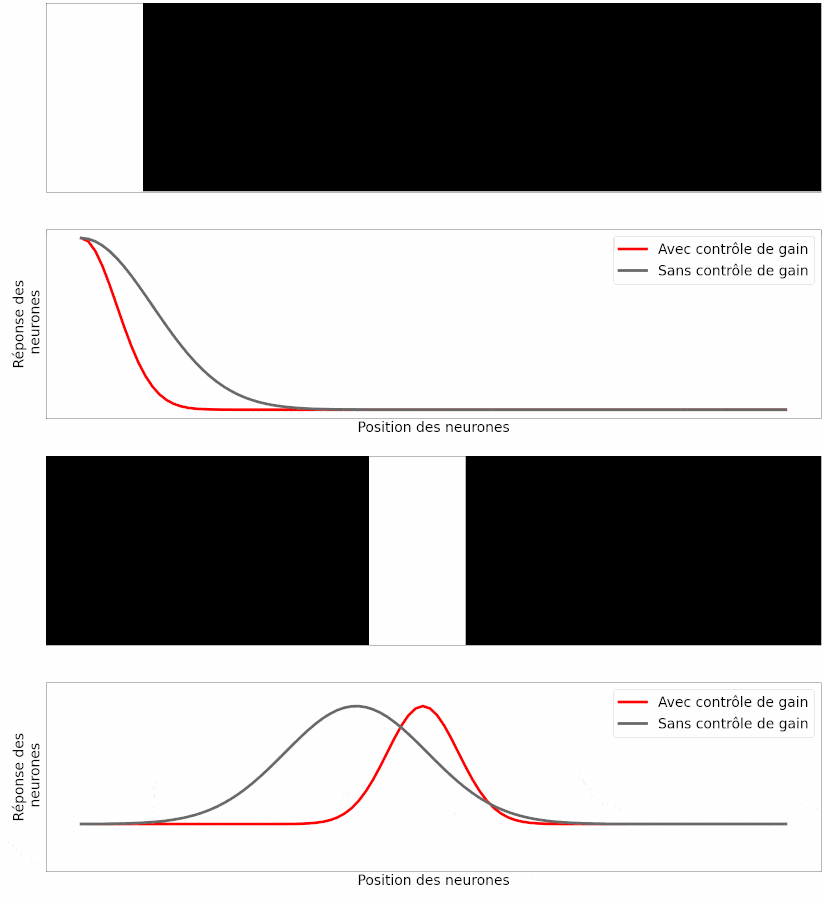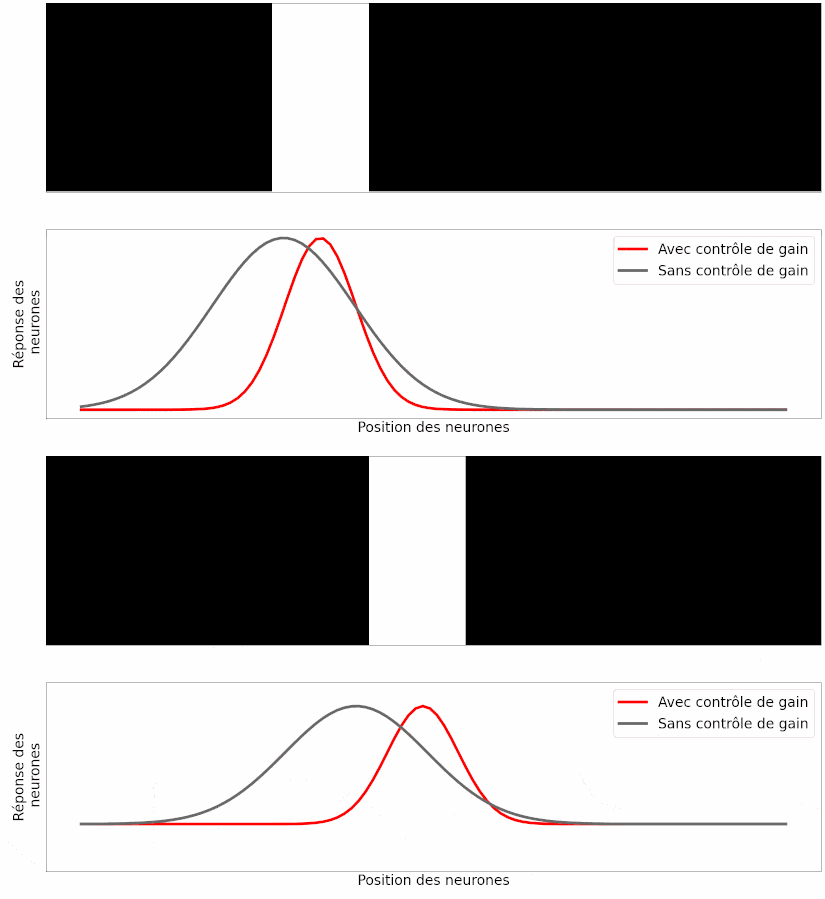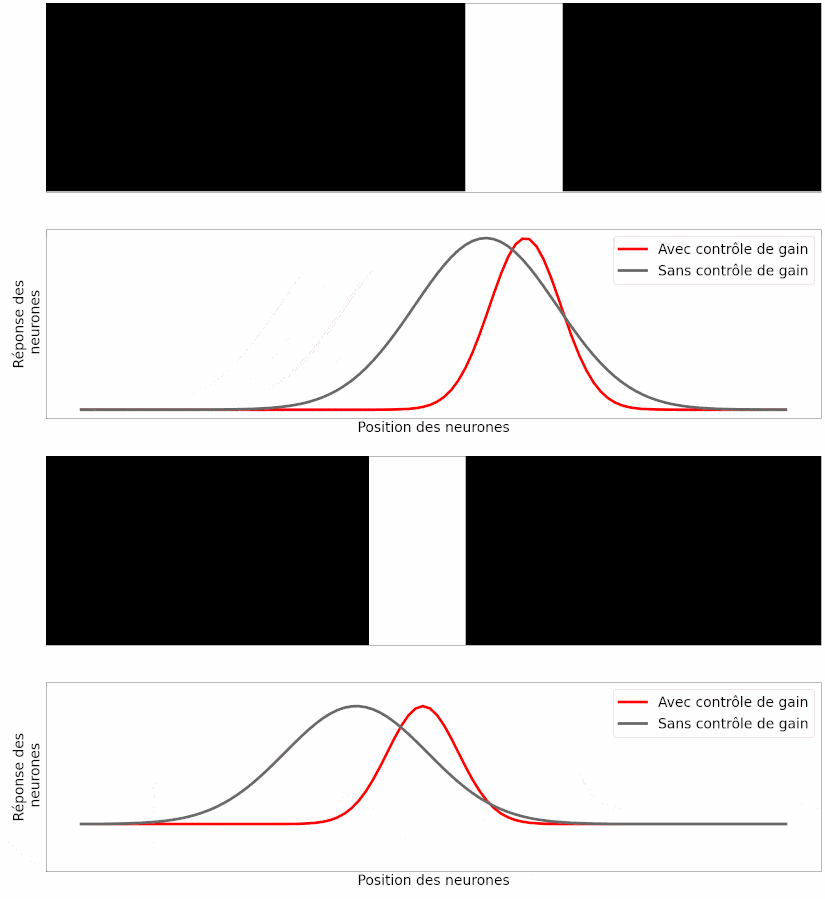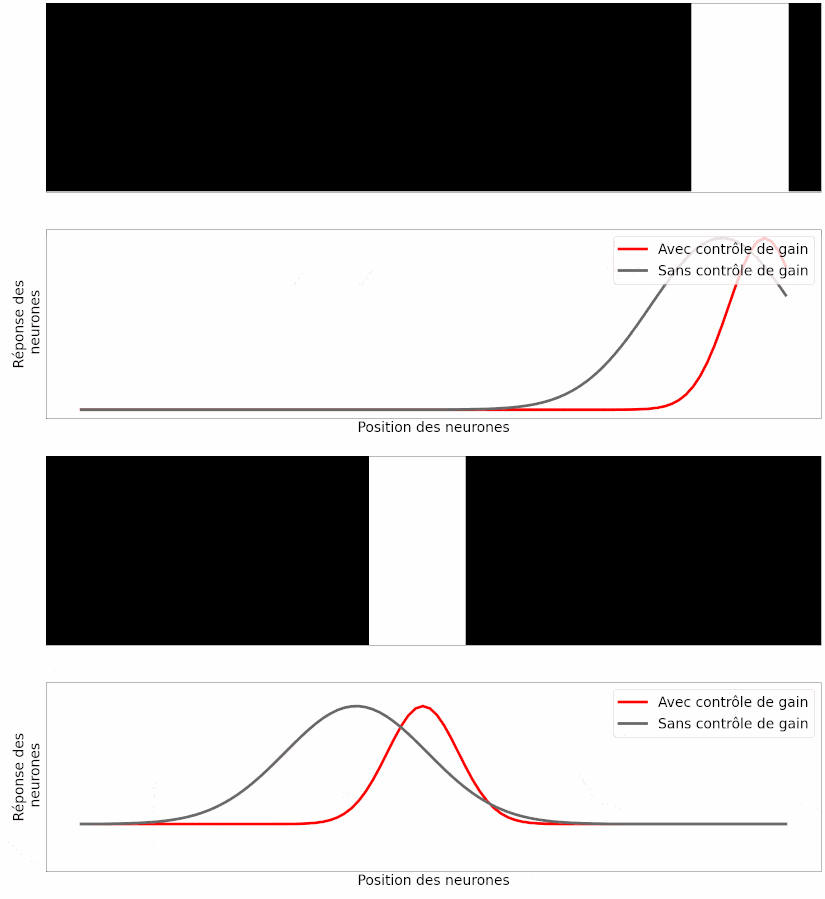
Le fait que les prédictions du modèle informatique correspondent aux résultats des expériences est un bon argument pour dire que ce mécanisme peut expliquer, au moins en partie, l’anticipation du mouvement [pour approfondir : voir note G].
Cette publication a donc permis de comprendre comment la rétine peut anticiper la trajectoire d’objets en mouvements. Un système ingénieux qui nous évite de vivre dans le passé, et qui peut même garder un dragon bien nourri.
Éléments pour approfondir
À partir des expériences d’électrophysiologie et de modélisation informatique, cet article a été le premier à décrire et expliquer un phénomène très particulier : l’anticipation du mouvement dans la rétine. Il présente plusieurs limites que nous allons détailler et qui sont tout à fait communes pour un article réalisant ce type de découverte. Mais surtout, cette publication a permis d’ouvrir de nouvelles pistes de recherche.
Note A
Modèles animaux. Les auteurs réalisent leurs expériences sur la rétine de lapin et de salamandre. Ce sont deux modèles animaux souvent utilisés dans l’étude du système visuel. En effet, ce sont tous deux des vertébrés, on peut donc espérer retrouver les résultats obtenus dans ces expériences chez d’autres espèces du même embranchement, notamment l’être humain. De plus, alors que le lapin est un mammifère, la salamandre est un amphibien, ce qui permet de justifier que les résultats obtenus sont bien conservés chez deux vertébrés assez éloignés phylogénétiquement, et qu’il ne s’agit donc pas d’un mécanisme très spécifique à une classe. Cependant, les résultats ne peuvent pas être extrapolés avec une absolue certitude chez d’autres espèces. Afin d’être certain que le même phénomène est présent, cette expérience doit être reproduite, comme cela a été fait depuis avec d’autres modèles animaux comme la souris et le poisson rouge [2].
Note B
Expériences ex vivo. Les expériences réalisées par les auteurs sont du type ex vivo. Cela signifie que l’animal a été euthanasié, que son œil a été disséqué et que l’activité des neurones a été enregistrée dans une boîte en verre contenant un milieu nutritif qui maintient la rétine vivante et fonctionnelle. Cette approche est utilisée principalement car elle simplifie énormément les expériences. Mais de la même manière que l’on ne peut pas extrapoler les résultats des expériences chez d’autres espèces, on ne peut pas être absolument sûrs que les résultats auraient été les mêmes chez l’animal vivant. Comme cette expérience n’a pas encore été réalisée, on n’a toujours pas la réponse à cette question.
Note C
Une stimulation visuelle artificielle. La stimulation visuelle utilisée par les auteurs, à savoir une barre lumineuse en mouvement sur un fond gris uniforme, est un standard dans le domaine des neurosciences de la vision. Elle a été utilisée pour étudier un grand nombre de phénomènes liés au mouvement, et pas seulement l’anticipation [3]. Mais elle ne correspond pas du tout à ce que verraient ces animaux dans leurs habitats naturels respectifs. Le phénomène d’anticipation du mouvement a donc été observé ici dans un contexte très particulier. Une tendance actuelle des neurosciences de la vision est d’utiliser des stimulations visuelles dites naturelles, c’est-à-dire plus proches des images vues par l’animal dans son quotidien. Des vidéos filmées dans les habitats des espèces étudiées sont par exemple utilisées [4]. Ces expériences apportent des résultats ayant plus de sens, mais sont plus difficiles à analyser à cause de la complexité des images utilisées, par rapport à un simple rectangle.
Note D
De l’anticipation, vraiment ? Les auteurs décrivent cette réponse particulière des neurones ganglionnaires à une barre en mouvement comme de l’anticipation. Ce terme évoque une prédiction de l’avenir, pourtant il permet uniquement aux neurones de transmettre une information actualisée. Un autre terme plus adéquat pour décrire ce phénomène pourrait être « compensation de délai ».
Note E
Quelques résultats supplémentaires. L’anticipation du mouvement de l’armée par les châteaux avec le nouveau système imaginé par la reine fonctionne uniquement pour une seule vitesse de l’armée : si elle se déplace de la distance séparant deux châteaux en un jour. De la même façon, le mécanisme proposé pour expliquer l’anticipation, le contrôle de gain, ne permet-il d’expliquer l’anticipation par les neurones de la rétine que pour une seule vitesse ? Pour répondre à cette question, les auteurs ont réalisé la même expérience en faisant cette fois varier la vitesse de la barre. Ils ont montré que l’anticipation était possible jusqu’à une certaine vitesse de l’objet, au-delà de laquelle le retard ne pouvait pas être compensé. L’activation des neurones dépend également fortement de la luminosité de la barre, ou plutôt de son contraste (différence de luminosité par rapport au fond). Pour tester l’importance de ce paramètre dans l’anticipation du mouvement, les auteurs ont répété la même expérience en faisant varier le contraste de la barre. Ils ont montré que l’anticipation diminuait lorsque le contraste de la barre est réduit.
Ces expériences supplémentaires ont donc montré que l’anticipation du mouvement fonctionne pour un objet en mouvement assez contrasté, et se déplaçant dans une certaine gamme de vitesse. Elles permettent d’une part de confirmer que l’anticipation observée précédemment n’était pas qu’un cas particulier, et d’autre part de tester les limites de cette anticipation. Elles apportent donc une meilleure caractérisation de l’effet et des observations supplémentaires pour tester le modèle, ce qui le rend plus fiable.
Note F
Une propriété émergente. Les résultats de cet article montrent qu’en modifiant uniquement le comportement individuel des éléments d’un système, ici les neurones ganglionnaires de la rétine, le système dans son ensemble peut acquérir des capacités intéressantes comme l’anticipation du mouvement. L’étude des relations entre les éléments d’un système et le système dans son ensemble est au cœur du domaine des neurosciences computationnelles, qui analyse les réseaux de neurones et les opérations que ceux-ci peuvent réaliser.
Note G
Validité du modèle. Dans cet article, l’effet observé de l’anticipation du mouvement est décrit précisément et est difficilement contestable. Les auteurs ont également exploré les limites de ce phénomène en faisant varier certains paramètres (vitesse et contraste de la barre). Cependant, le mécanisme permettant cette anticipation peut être discuté. Le raisonnement que les auteurs utilisent pour l’argumenter est courant dans le domaine des neurosciences computationnelles et est le suivant. La rétine est un réseau avec une certaine structure, un certain fonctionnement et un certain comportement. On est capable de réaliser des modèles informatiques de réseaux qui ont une structure et un fonctionnement comparables à ceux d’une « vraie » rétine. Si le réseau modélisé a un comportement similaire à la vraie rétine, on suppose alors que la structure et le fonctionnement de la vraie rétine, qui sont la cause du comportement, sont comparables à ceux du modèle. Mais cela n’est pas nécessairement vrai. Le modèle peut être une façon parmi d’autres d’arriver au résultat, sans être nécessairement celle qui est utilisée par la vraie rétine.
Pour utiliser une analogie, c’est comme essayer de deviner le moyen de transport d’une personne à partir de son temps de trajet. Imaginons que l’on sache qu’une personne a mis 5 heures pour aller de Paris jusqu’à Lyon. En réalisant une simulation sur un site de cartographie, on constate que le trajet Paris-Lyon prend 5 heures en TER. On voit que notre hypothèse, un trajet en TER, donne le même résultat que la réalité : un trajet en 5 heures. On peut donc faire l’hypothèse que cette personne s’est déplacée en TER. Mais si on simule le temps de trajet en voiture, on constate qu’il dure également 5 heures. L’hypothèse d’un trajet en voiture est également valable en suivant le même raisonnement ! Plusieurs chemins permettent d’arriver à la même destination, et on ne peut pas décider, uniquement à partir de cette unique observation, lequel est le bon. De la même manière, la correspondance du modèle avec les données de l’expérience est un argument pour dire qu’il est correct, mais on ne peut pas conclure définitivement.
[1] Baldo M. V. C. & Klein S. A., Extrapolation or attention shift? Nature, 1995. DOI : 10.1038/378565a0. [Publication scientifique]
[2] Johnston J., et al., A retinal circuit generating a dynamic predictive code for oriented features. Neuron, 2019. DOI : 10.1016/j.neuron.2019.04.002. [Publication scientifique]
[3] Euler T., et al., Directionally selective calcium signals in dendrites of starburst amacrine cells. Nature, 2002. DOI : 10.1038/nature00931. [Publication scientifique]
[4] Qiu Y, et al., Mouse retinal specializations reflect knowledge of natural environment statistics. bioRχiv, 2020. DOI : 10.1101/2020.12.08.416172. [Prépublication, article non relu par les pairs]
Publié le 12/06/2021
 Thomas Buffet et Pouaseuille/Papier-Mâché/CC BY-NC-SA 4.0 2021
Thomas Buffet et Pouaseuille/Papier-Mâché/CC BY-NC-SA 4.0 2021Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.