Chargement de l'article...
Utilisation des corrélats immunitaires pour comprendre la progression du SIDA
Écriture : Aurélien Schwob
Relecture scientifique : Lise Chauveau et Paul Verlhac
Relecture de forme : Félicités Rapon et Eléonore Pérès
Temps de lecture : environ 20 minutes.
Thématiques : Immunologie et Microbiologie (Biologie)
Publication originale : Cumont MC., et al., Early divergence in lymphoid tissue apoptosis between pathogenic and nonpathogenic simian immunodeficiency virus infections of nonhuman primates. Journal of Virology, 2007. DOI : 10.1128/jvi.00450-07. Accès libre sur HAL (pdf).

Que ce soit pour résoudre un exercice complexe de géométrie, apprendre à pêcher ou à jouer à la belote, qui n’a jamais cherché de l’aide auprès d’une personne de sa famille ? Certains scientifiques ont poussé l’idée un peu plus loin pour tenter d’élucider certains mystères qui persistent autour de la pathologie du SIDA, en étudiant l’expérience de nos cousins les singes, qui ont un long vécu avec des virus proches. Loin d’une paisible partie de pêche, cette aventure pourrait bien remettre en cause ce que vous croyez savoir sur les tenants et aboutissants de cette maladie…
On attribue généralement à Confucius la maxime suivante : « L’homme sage apprend de ses erreurs, l’homme plus sage apprend des erreurs des autres ». Ce proverbe qui nous incite à profiter des expériences et enseignements de nos pairs, nous l’appliquons consciemment ou non toutes et tous au long de nos parcours de vie. C’est d’ailleurs une caractéristique humaine à l’échelle de l’évolution. En effet, notre espèce est notamment caractérisée par une longue phase juvénile au cours de laquelle les jeunes apprennent au contact de leur entourage. Plus précisément, cette caractéristique est commune à l’ensemble du groupe des primates. L’apprentissage joue un rôle primordial chez tous les singes, qu’ils soient humains ou non !
Au cours de cet article, nous allons voir comment prendre au mot cette maxime pour engranger des connaissances sur un thème particulièrement pointu : celui des agents pathogènes et des maladies infectieuses. En effet, les interactions entre les micro-organismes et leurs hôtes sont d’une grande complexité. Parasites mortels, compagnie insignifiante ou partenaires utiles, on trouve tous les types d’interactions entre ces petites bestioles et notre organisme, chaque cas étant unique et résultant d’une longue histoire évolutive.
Mais lorsque nous sommes confrontés à un pathogène actuel, il est impossible d’avoir une vision claire sur son évolution, et donc de la relation qu’il entretient avec son hôte. Cet état de fait frustrant peut être contourné grâce à nos connaissances sur les mécanismes de l’évolution. En effet, les liens de parenté évolutifs qui unissent les espèces ne s’arrêtent pas aux hôtes des agents pathogènes (par exemple entre un singe et un humain) mais ont aussi cours entre les pathogènes eux-mêmes. Afin d’éclaircir certains aspects des relations que nous entretenons avec nos propres pathogènes, il est donc possible d’étudier les interactions entre nos cousins les singes et leurs pathogènes. En effet, comme nous sommes nous-mêmes proches des singes, qui appartiennent à la même grande famille d’espèces que nous (celle des primates), les pathogènes des singes sont eux-mêmes de proches cousins de nos propres pathogènes.
Une équipe française de l’INSERM s’est lancée dans cette démarche, en étudiant certains singes qui réagissent de diverses manières à l’infection par des virus proches du point de vue de l’évolution du Virus de l’Immunodéficience Humaine (VIH). Notre expérience humaine nous dicte que toutes les personnes infectées par le VIH vont développer, à plus ou moins long terme, une pathologie appelée Syndrome d’ImmunoDéficience Acquise (SIDA). Pourtant, après infection par leurs équivalents simiens (chez les singes ce sont les Virus de l’Immunodéficience Simienne ou VIS), certains de nos proches cousins ne développent pas de pathologie particulière. Il nous est donc possible, par comparaison, d’identifier les facteurs qui favorisent, chez nous/l’humain, la progression vers la pathologie du SIDA.
Étudier les facteurs de la progression vers le SIDA : le serpent qui se mord la queue
Le Syndrome d’ImmunoDéficience Acquise, ou SIDA, est une maladie mortelle touchant l’espèce humaine. Elle est caractérisée par une phase d’immunodéficience, c’est-à-dire un état dans lequel notre système immunitaire n’est plus capable de défendre l’organisme contre les menaces externes (infections) ou internes (cancers). Lors de cette phase, les patients concernés développent des maladies dites opportunistes, c’est-à-dire des pathologies normalement très bien contrôlées par le système immunitaire et qui ne touchent donc que les immunodéprimés. Touchés par ces maladies opportunistes (comme divers cancers) et plus exposés aux autres pathogènes (virus ou autres bactéries), les patients décèdent précocement s’ils n’ont pas accès à des traitements adaptés.
Pour comprendre cette immunodéficience, penchons-nous sur le fonctionnement du système immunitaire, et plus précisément sur une de ses réponses appelée réponse adaptative, qui est la réponse la plus efficace de l’immunité humaine. Cette réponse fait intervenir 3 types de cellules de la famille des lymphocytes, que l’on distingue par certaines molécules qu’elles portent à leur surface. Les lymphocytes T portent un récepteur appelé le récepteur T, ainsi qu’un co-récepteur qui peut être le CD4 ou le CD8. Cela nous permet de définir les lymphocytes T CD4 et les lymphocytes T CD8. D’autres lymphocytes portent le récepteur B : ce sont les lymphocytes B.
Lors de l’activation de la réponse immunitaire, un lymphocyte T CD4 est activé suite à la détection du pathogène. Ce lymphocyte T CD4 est le chef d’orchestre de cette réponse adaptative, et il va à son tour aider l’activation des lymphocytes T CD8 et des lymphocytes B. Les lymphocytes T CD8 se différencient (se transforment) en lymphocytes capables de détruire les cellules infectées par des virus notamment : on les appelle des lymphocytes T cytotoxiques. Les lymphocytes B se différencient quant à eux en cellules capables de produire des anticorps qui vont permettre de lutter contre les pathogènes, on les appelle des plasmocytes (Figure 1).
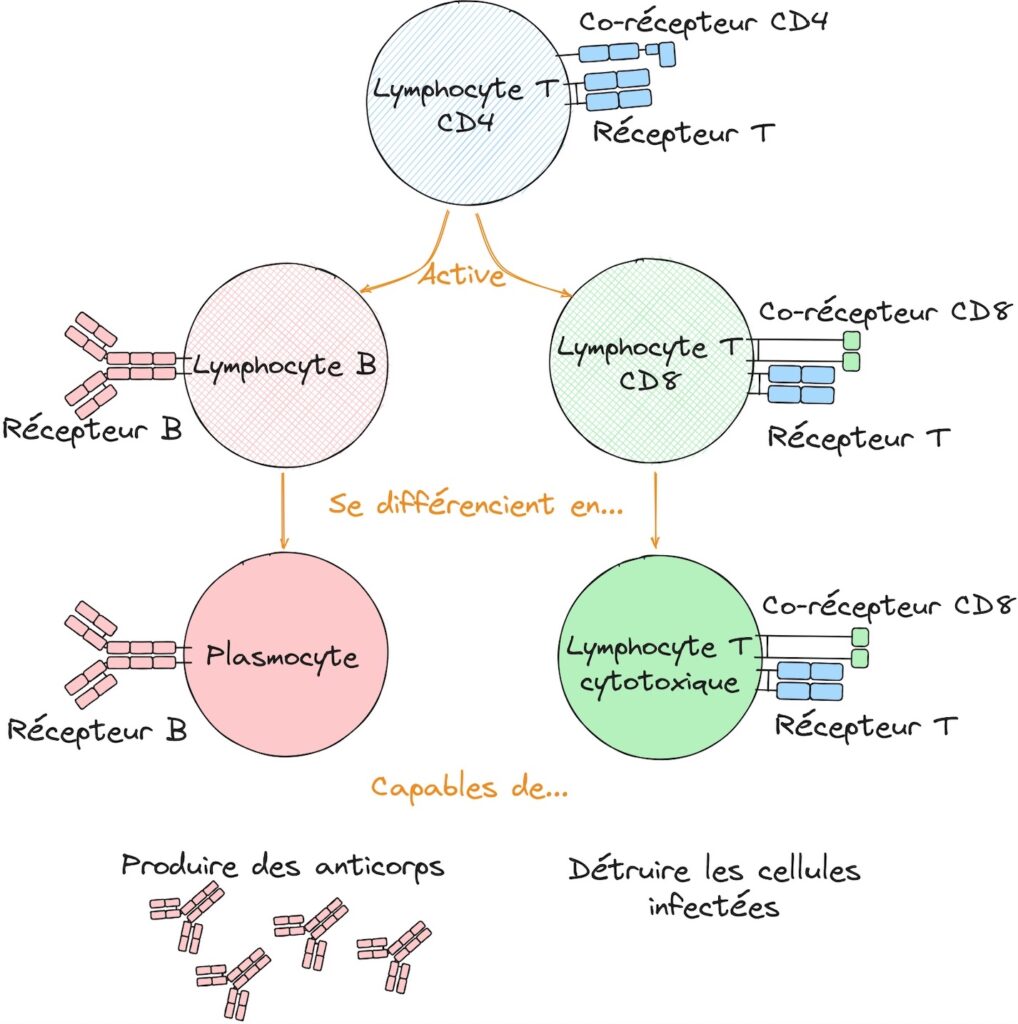
Or, dans le cas du SIDA, ce sont les lymphocytes T CD4 qui perdent leur fonction. Ainsi incapables de coordonner la réponse immunitaire, cette dernière devient inefficace et le corps ne peut lutter contre les maladies opportunistes. On sait depuis la découverte de Françoise Barré-Sinoussi et Jean-Claude Chermann en 1982 que le SIDA est provoqué par l’infection par un virus appelé le Virus de l’Immunodéficience Humaine [1]. Ce virus infecte notamment les lymphocytes T CD4 et la multiplication du virus dans ces cellules provoque des dysfonctionnements et leur destruction [2]. Vu d’ici, on pourrait se dire que le mécanisme est simple : le VIH va infecter un à un tous les lymphocytes T CD4 jusqu’à épuisement, amenant à leur destruction totale et donc à l’arrêt de l’immunité. Mais en réalité il n’est pas si clair que ce mécanisme soit suffisant pour expliquer l’épuisement des lymphocytes. En effet, au fil des ans, d’autres hypothèses ont émergé comme une destruction indirecte de ces lymphocytes, une simple inactivation ou encore le résultat d’une réponse immunitaire trop élevée ou au contraire, trop faible (Figure 2).
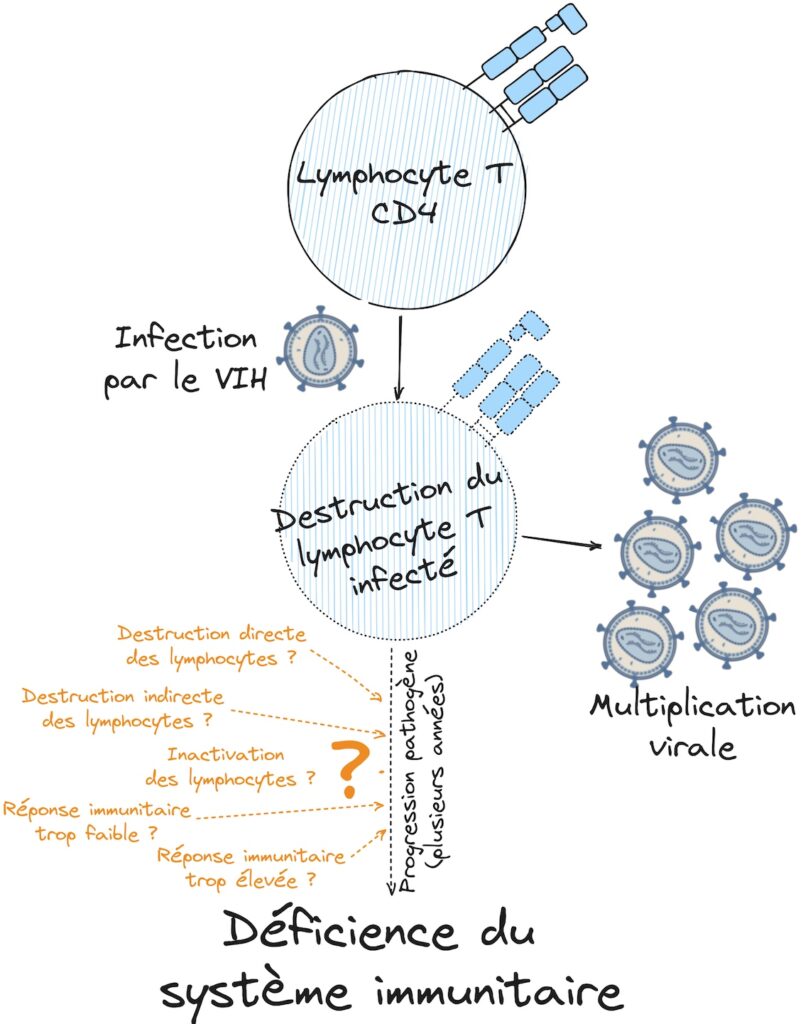
Mais le problème, c’est qu’il est très difficile de tirer cette affaire au clair. En effet, les lymphocytes étant à la fois cibles du VIH et acteurs de la destruction des cellules infectées, comment interpréter les modifications qui affectent ces lymphocytes ? Est-ce un effet du pathogène sur leurs cibles ? ou bien un effet de l’action des lymphocytes dans la lutte contre les pathogènes ? D’ailleurs, parmi tous les patients progressant jusqu’à la phase SIDA (plus ou moins vite), on ne peut pas faire de comparaisons simples entre un cas pathologique et un cas non pathologique. Pire, même si on pouvait faire ces comparaisons, on ne pourrait savoir quel patient progresse rapidement ou lentement vers le SIDA que des années après l’infection, sans avoir la possibilité d’étudier la réaction de son système immunitaire initialement !
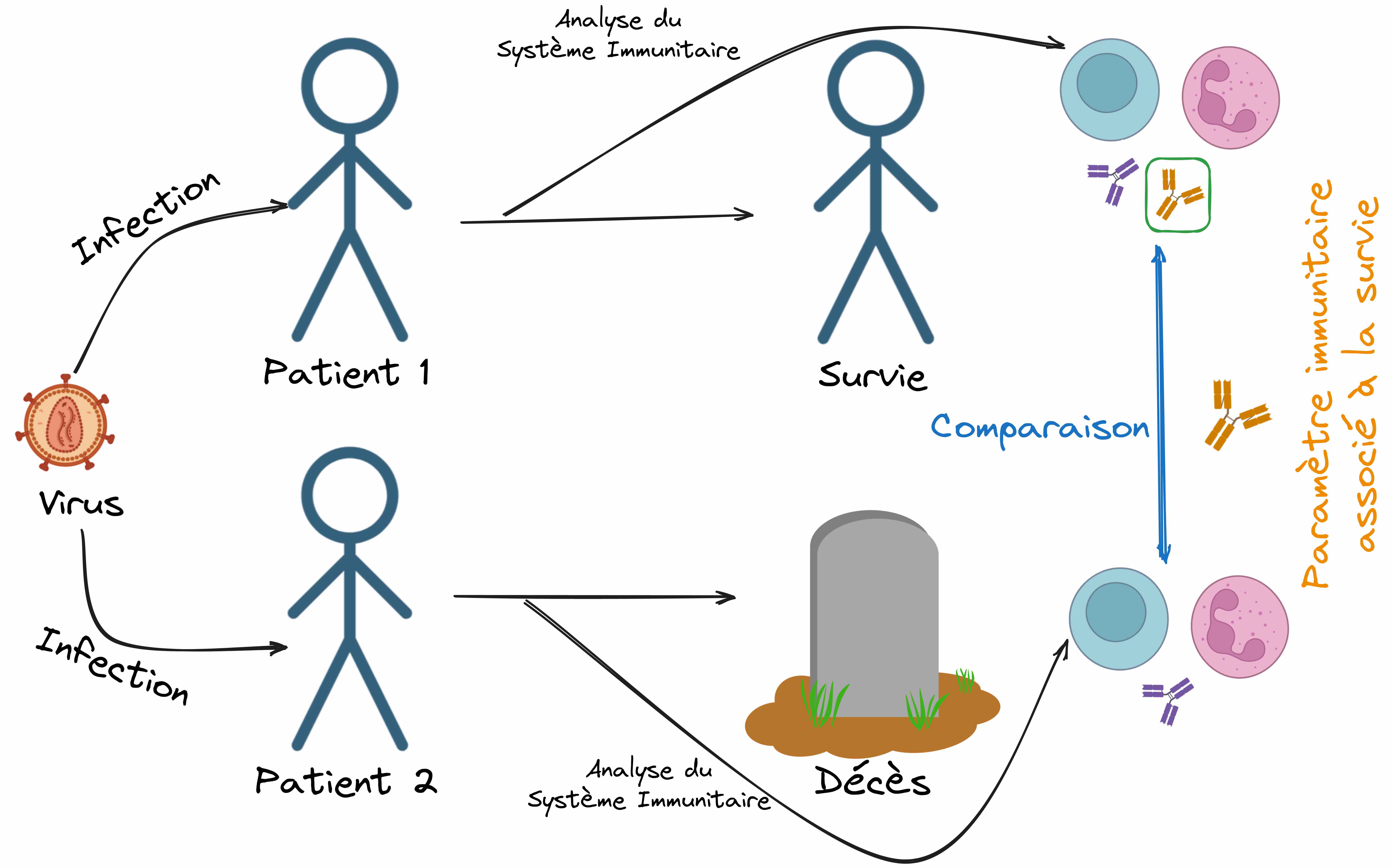
C’est dans ce contexte que la communauté scientifique spécialiste de la question s’est tournée vers des outils puissants. D’une part, la comparaison entre les cas humains et simiens (les autres singes). Ensuite, l’exploitation de ce que l’on appelle des « corrélats immunologiques », à savoir l’identification de mécanismes immunitaires qui sont associés à la gravité de la pathologie telle qu’elle se développe réellement chez le patient et aident à l’expliquer. Pour illustrer avec un exemple un peu caricatural, considérons deux patients infectés par le même virus, mais dont l’un survit et le second décède. Si, après analyse de leur système immunitaire, on se rend compte que le patient ayant survécu a produit des anticorps spécifiques de ce virus et que le patient ayant trépassé n’a pas produit d’anticorps, on aura identifié un mécanisme immunitaire associé à la progression de la pathologie pour ce virus (Figure 3).
Des primates qui ne se contentent pas de singer l’infection humaine
En effet, nos cousins évolutifs primates possèdent eux aussi des équivalents du VIH, que l’on appelle les VIS (qui pour rappel signifie Virus de l’Immunodéficience Simienne). Mais comme leur histoire évolutive avec ces pathogènes est par définition différente (puisqu’il n’y a aucune raison pour que, au cours de l’évolution, se produisent exactement les mêmes événements à partir d’une même situation initiale), leur réaction l’est probablement aussi.
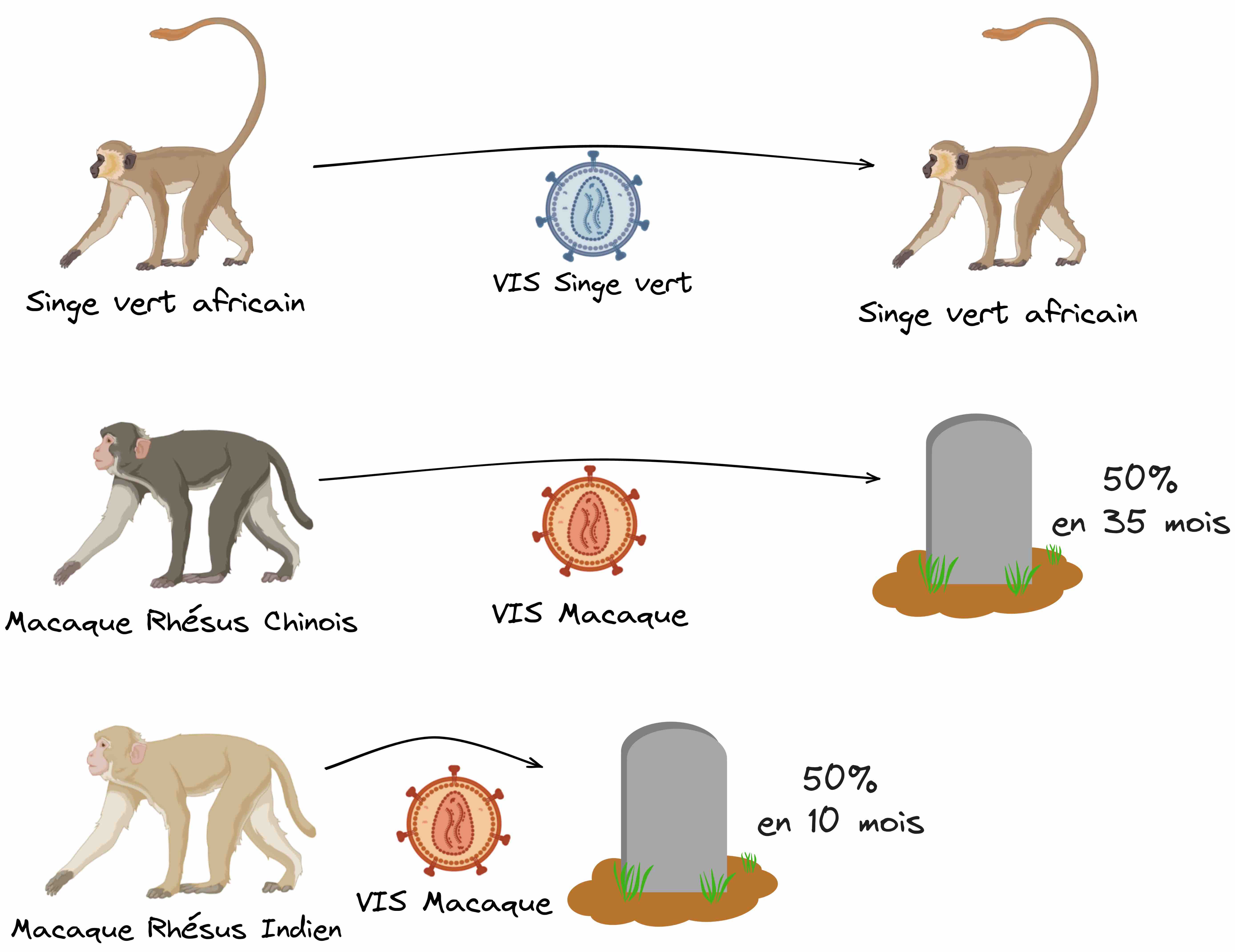
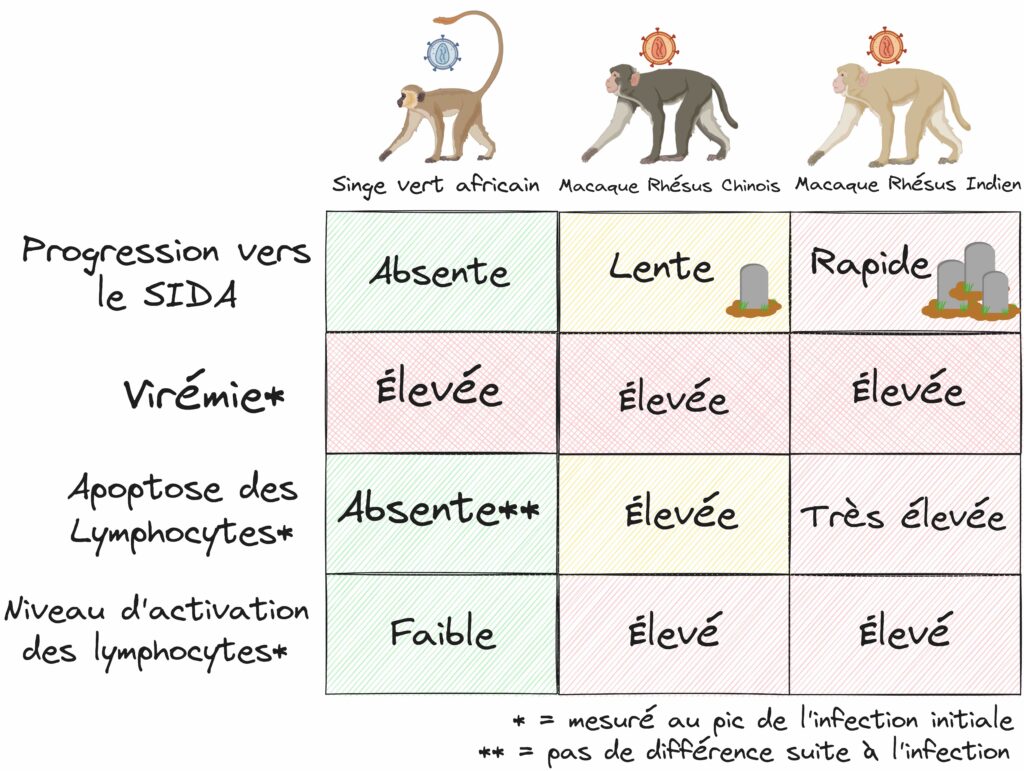
Prenons par exemple l’espèce du Macaque Rhésus. Cette espèce vivant en Asie du Sud-Est peut être infectée par un VIS spécifique (le VIS macaque), ce qui provoque systématiquement une progression vers une phase SIDA et le décès des animaux infectés, exactement comme chez l’hôte humain. À l’inverse, chez le Singe Vert Africain, l’infection par son VIS spécifique (le VIS singe vert) ne provoque pas de pathologie. On a ainsi un modèle comparatif entre des infections pathologiques et non pathologiques, permettant de savoir quels sont les facteurs caractéristiques de la progression vers la pathologie. C’est cette méthode qu’a utilisée en 2007 une équipe de recherche. Mieux encore, pour modéliser les différents rythmes de progression vers la phase SIDA, cette équipe a introduit dans son étude comparative deux sous-populations de Macaques Rhésus : une population chinoise, qui progresse lentement vers le SIDA (la moitié des individus décède 35 mois après infection) et une population indienne qui progresse rapidement vers le SIDA (la moitié des individus est ici décimée en à peine 10 mois) (Figure 4).
Après avoir infecté des populations de Singe Vert Africain avec le virus VIS singe vert et une population de Macaques Rhésus de Chine ainsi qu’une population de Macaque Rhésus d’Inde toutes les deux avec la même souche de VIS macaque [voir la version Approfondissement pour plus d’informations sur le choix de cette souche], l’équipe a mesuré un certain nombre de paramètres sur ces animaux. Une fois les différences de progressions pathologiques exposées plus haut (Figure 4) vérifiées, la première chose à faire a été de mesurer la virémie. La virémie désigne la quantité de virus circulant dans le sang. L’infection par les VIS, comme le VIH, se caractérise par une première phase dite aiguë où le virus se réplique beaucoup, suivie par une phase dite chronique où la virémie baisse. Les chercheuses et chercheurs ont donc mesuré principalement la virémie à son pic d’infection initiale. Cela permet également de chercher les événements précoces prédictifs d’une progression pathogénique plus ou moins rapide.
Dans la logique de l’hypothèse selon laquelle la destruction des lymphocytes est le résultat de la seule infection virale qui détruirait les cellules une à une, on s’attend à ce que la virémie soit proportionnelle à la pathologie : plus les virus infectent de cellules, plus ils se multiplient et plus les cellules immunitaires sont détruites (Figure 5).
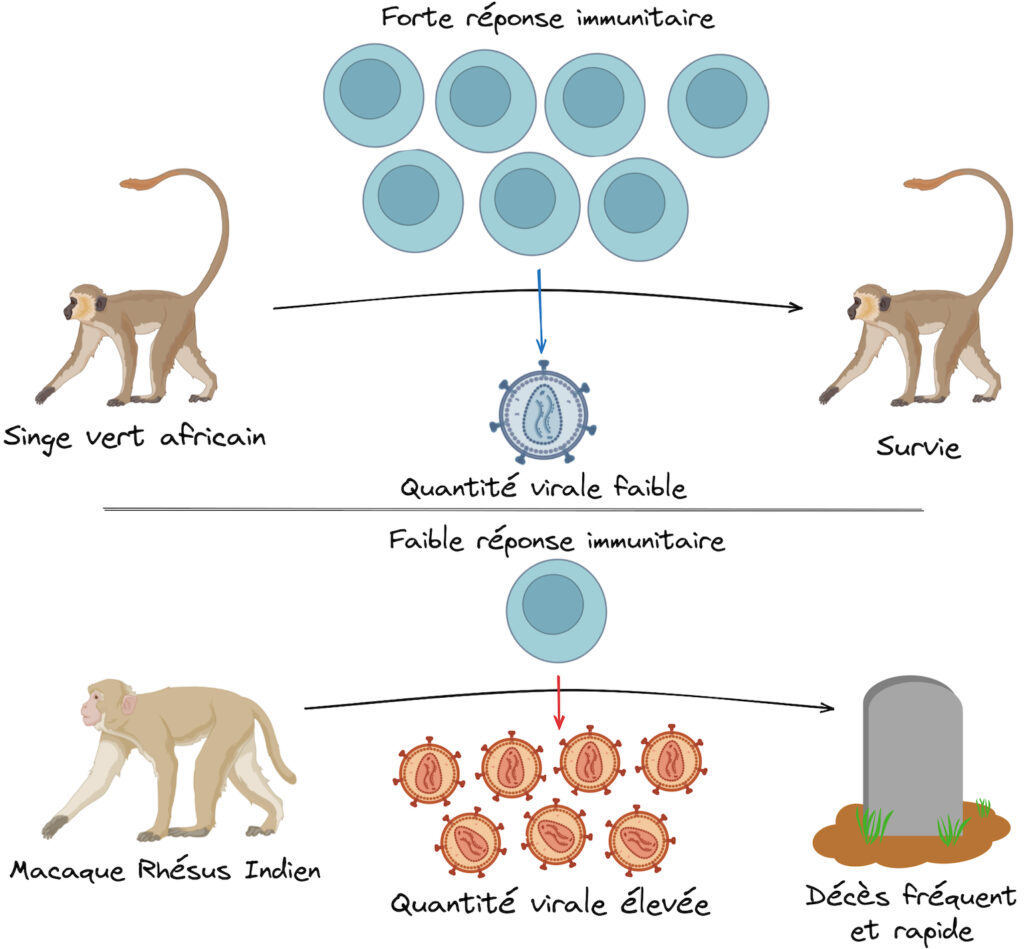
Dans cette hypothèse, le SIDA serait provoqué par une réponse immunitaire trop faible pour contrer le virus. Or, lorsque l’équipe de recherche compare la virémie au pic de l’infection entre les trois conditions étudiées, ils observent que cette virémie n’est pas significativement différente entre elles ! En effet, que ce soit chez le Singe Vert Africain ne développant pas de pathologie, le Macaque Rhésus Chinois progressant lentement vers le SIDA ou le Macaque Rhésus Indien progressant rapidement vers le SIDA, on trouve toujours entre approximativement 1 million et 100 millions de virus par millilitre de sang ! Ces résultats, conformes à d’autres observations déjà réalisées à l’époque, portent un sérieux coup à l’hypothèse d’une destruction « manuelle » des cellules immunitaires par le virus… et rendent caduque l’idée de prédire la gravité de la progression pathologique en se basant sur la virémie seule.
Des singes qui permettent de lever un lièvre : le rôle de l’apoptose des lymphocytes T
Après cette première observation pouvant sembler étonnante, l’équipe de recherche a poussé plus loin ses investigations. Une hypothèse pour expliquer l’absence des lymphocytes T est que leur destruction se fasse par un processus appelé apoptose. L’apoptose, ou « mort cellulaire programmée » est un phénomène biologique intégré au fonctionnement normal de l’organisme par lequel les cellules vont fragmenter leur ADN et se détruire « de l’intérieur », ne laissant derrière elles que des petites vésicules contenant les débris cellulaires qui seront ensuite « nettoyés » par des cellules immunitaires.
Ce phénomène a lieu à plusieurs étapes du développement de l’organisme, de manière tout à fait normale et contrôlée. C’est par exemple par apoptose que les cellules de la peau située entre nos doigts sont détruites lors du développement embryonnaire, nous permettant d’arborer des mains non palmées. C’est également l’apoptose qui permet la destruction des cellules trop vieilles ou qui ont accumulé des dommages cellulaires. C’est encore par apoptose que les lymphocytes qui ciblent des éléments de notre propre organisme sont éliminés lors de leur formation, ou encore que la plupart des nombreux lymphocytes produits en réponse à une infection sont détruits à la fin de cette infection. Ces deux derniers exemples nous montrent que l’apoptose concerne aussi les cellules immunitaires. Il se pourrait donc que le virus provoque, directement ou indirectement, la mort de ces cellules par ce phénomène d’apoptose.
Les scientifiques ont donc quantifié le phénomène d’apoptose à l’intérieur de ganglions lymphatiques, ces structures qui enflent lorsque nous sommes infectés et qui sont le lieu de rencontre et de multiplication des cellules immunitaires suite à la détection d’un pathogène. Par une technique un peu complexe mais assez visuelle permettant de colorer les cellules dont l’ADN est fragmenté, l’équipe de recherche a visualisé directement au microscope le nombre de cellules en apoptose dans les ganglions prélevés chez les différents animaux infectés. Les résultats montrent que l’apoptose augmente fortement au pic de l’infection chez les Macaques Rhésus Chinois, encore un peu plus chez les Macaques Rhésus Indiens alors que chez le Singe Vert Africain, l’apoptose n’augmente pas suite à l’infection. Confirmant des suppositions précédentes, l’équipe vient ainsi de mettre la main sur un facteur qui semble proportionnel à la progression pathogénique ! L’apoptose des lymphocytes au pic de l’infection (donc lors des phases précoces) est proportionnelle à la rapidité de la progression pathogénique. Si cela semble assez logique, il convient de noter plusieurs éléments une fois cette conclusion faite. Le fait que le taux d’apoptose soit décorrélé de la virémie montre bien que cette hausse de l’apoptose n’est pas directement liée au virus, et qu’il existe d’autres mécanismes au niveau cellulaire expliquant la progression vers la phase SIDA.
Quand le système immunitaire monte sur ses grands chevaux
C’est vers l’origine de cette hausse de l’apoptose que nous allons nous tourner pour finir notre exploration de certains résultats de cette publication. En effet, nous l’avons dit précédemment, l’apoptose est un phénomène normal concernant les lymphocytes en fin d’infection ou lorsque l’infection devient chronique. Lorsque vous êtes infectés par un virus chronique comme certains virus de la famille des Herpèsvirus par exemple (comme les virus à l’origine des boutons de fièvre ou des crises d’herpès), vous allez subir une phase d’infection aiguë accompagnée de symptômes et d’une grande activation de lymphocytes. Mais après quelques jours, la quantité de virus baisse sans que ces derniers n’aient été éradiqués, et la quantité de lymphocytes baisse également : c’est la phase chronique. On pourrait donc chercher à corréler cette forte apoptose avec un fort taux d’activation immunitaire.
C’est notamment ce qu’ont étudié les scientifiques de l’équipe que nous suivons [parmi d’autres paramètres ; voir la version Approfondissement pour plus en savoir plus]. Ils et elles ont cherché à estimer le niveau d’activation des lymphocytes entre nos trois conditions. Pour cela, c’est assez simple : vous vous souvenez du fait que les récepteurs à la surface des cellules nous renseignent sur leur nature (lymphocytes T CD4, T CD8 et B) ? Et bien certains récepteurs nous renseignent de manière plus fine sur l’état d’activation des cellules. En ciblant les récepteurs qui sont connus pour marquer une activation des cellules, il est possible de marquer avec une molécule fluorescente ces cellules. Faites passer vos cellules ainsi étiquetées « actives » ou « inactives » devant un lecteur automatisé et vous obtiendrez le taux de cellules actives, et ainsi le niveau d’activation globale de vos lymphocytes. En utilisant cette méthode (ce qui est, vous l’imaginez, presque aussi simple à dire qu’à faire, et j’ai bien dit presque), les chercheurs ont fait une autre découverte intéressante.
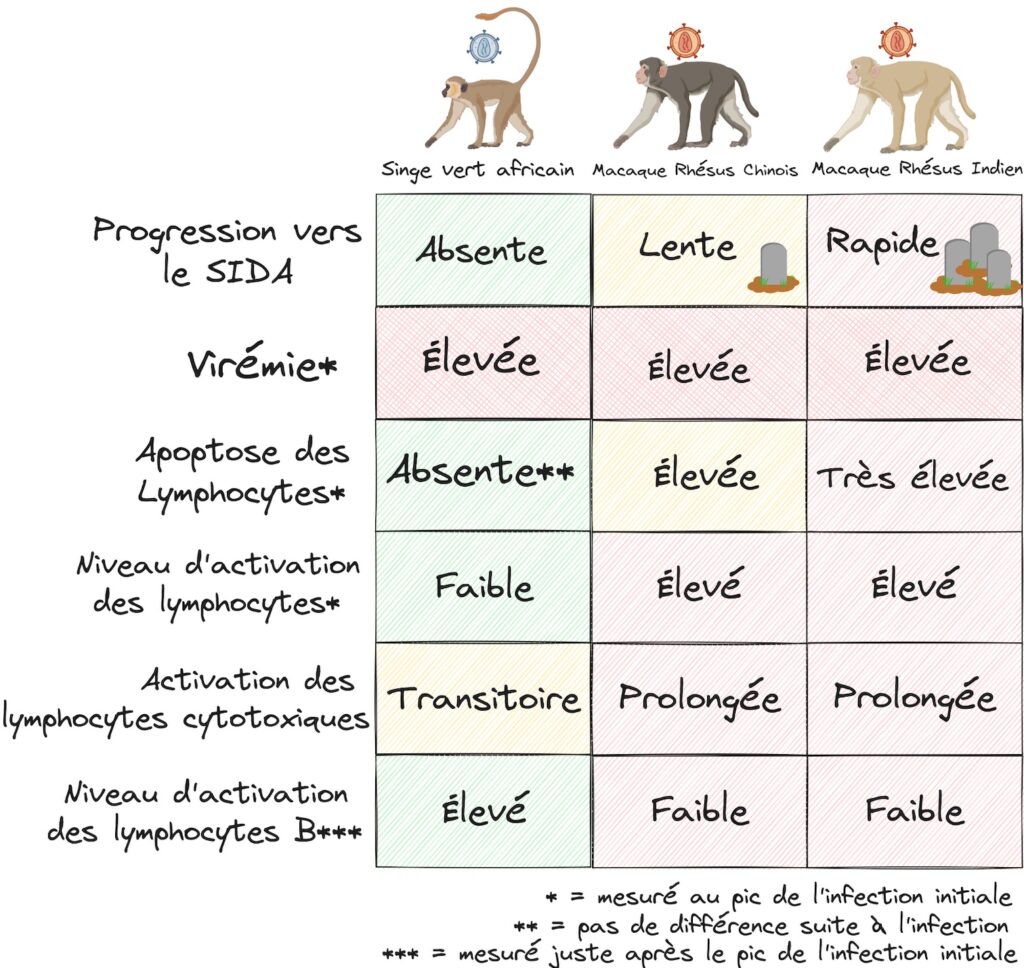
En effet, lorsque l’on compare le niveau d’activation des lymphocytes entre nos trois types de singes, on observe tout d’abord que les deux Macaques Rhésus sont caractérisés par un niveau d’activation des lymphocytes bien plus élevés que le Singe Vert Africain (avec environ 4 fois plus de cellules actives au pic infectieux). Cela confirme que la faible apoptose des singes ne développant pas de pathologie SIDA (le Singe Vert Africain) est corrélée à un faible taux d’activation des lymphocytes par rapport aux singes développant une pathologie SIDA. Mais plus intéressant encore, on ne trouve pas de différence significative entre les deux populations de Macaque Rhésus, alors même qu’il y a une différence de progression pathologique ! Cela montre que cette approche de corrélats immunologiques chez les primates non-humains permet non seulement d’établir des corrélations entre les événements précoces de l’infection et la gravité de la progression pathologique, mais qu’en plus cette méthode permet de distinguer les effets liés au virus et les effets liés à l’hôte infecté. En effet, les Macaques Rhésus Chinois ont une apoptose plus faible que les Macaques Rhésus Indiens, malgré un niveau d’activation similaire des lymphocytes et le fait qu’ils sont infectés par le même virus (Figure 6). Cela montre qu’entre ces deux populations, c’est bien une différence au niveau de la manière dont leurs organismes réagissent à un même virus qui conditionne la pathologie. Écho à l’idée plus générale que le caractère pathologique d’une infection d’un hôte par un agent infectieux dépend en grande partie de leur histoire évolutive commune, et de leurs possibles adaptations au cours du temps.
Conclusion : les singes qui murmurent à l’oreille des scientifiques
À l’aide de corrélats immunitaires, cette étude a donc approfondi nos connaissances dans la pathogenèse du VIH et sa progression jusqu’à la déclaration de la phase SIDA. En partant des données déjà disponibles sur les différences de vitesse de progression vers le SIDA de plusieurs espèces simiennes et sur l’idée déjà connue d’un potentiel lien entre cette progression et l’apoptose des lymphocytes T, les scientifiques à l’origine de cette étude ont permis d’élucider les mécanismes qui sous-tendent l’apoptose des lymphocytes T, permettant de faire progresser la question de l’origine de la phase SIDA lors d’une infection par un VIS.
En effet, en démontrant le lien entre apoptose des lymphocytes T et leur niveau d’activation, ces travaux ont participé à la démonstration que la pathogenèse du SIDA était liée, de manière contre-intuitive, à un excès de réponse immunitaire des organismes face au VIS. Les espèces qui ne développent pas de pathologie suite à l’infection ne sont pas particulièrement capables d’éradiquer le virus, mais sont caractérisées par une réponse immunitaire initiale moins forte. Pour simplifier, cette suractivation du système immunitaire va provoquer in fine son épuisement, là où une réponse plus modérée permet de sauvegarder les forces du système immunitaire. Et sans l’apport de nos cousins primates non-humains, difficile d’imaginer que le problème à l’origine d’une maladie caractérisée par une immunodéficience réside dans le fait d’avoir une réaction immunitaire trop forte !
Cette idée a depuis fait son chemin et fait maintenant consensus, à tel point que des pistes thérapeutiques sont envisagées consistant à modérer la réponse immunitaire des patients pour ralentir la progression SIDA [3].
Tout cela met en évidence la puissance de l’utilisation des corrélats immunitaires. Nous en avons ici un exemple utilisant des espèces non-humaines, mais de manière générale cette méthode a de nombreuses applications. Plutôt que d’essayer de comprendre les mécanismes sous-jacents dans un système d’interactions éminemment complexe, l’étude des corrélats se veut plus statistique. Prenons deux types de patients soumis au même agent pathogène, l’un développant une pathologie rapide et l’autre montrant peu de symptômes. En comparant leurs réponses immunitaires, cela nous donnera autant de pistes de compréhension des mécanismes à l’œuvre, et même, des pistes thérapeutiques !
Bien entendu, les conclusions de cette étude seule ne doivent pas nous emmener trop loin dans notre enthousiasme. Aussi brillante que soit l’idée de comparer nos cousins évolutifs infectés par les cousins évolutifs de nos virus, il faut garder en tête que des différences importantes peuvent exister entre ces espèces animales et virales. Ainsi, il serait possible que ces conclusions valables pour les interactions singes / VIS ne puissent s’appliquer au contexte de l’hôte humain / VIH. Cette réserve a donc nécessité des investigations complémentaires visant à confirmer ces hypothèses pour les hôtes humains. Mais la forte parenté entre primates non-humains et humains, ainsi que la parenté particulièrement forte entre VIS et VIH (le second dérivant récemment du premier [4]) permet d’accorder beaucoup de crédit à ces découvertes dans ce contexte précis.
Au final, en plus de nous apporter des réponses sur la pathogenèse des VIS et VIH, cette étude nous rappelle que lorsqu’il s’agit d’interactions hôtes/pathogènes, tout est question d’histoire évolutive commune et d’adaptation des organismes les uns aux autres. Cela montre encore une fois la place centrale des concepts évolutifs dans toutes les branches de la biologie. Reste à savoir si, comme Confucius nous y incite, ces découvertes nous permettront d’en apprendre assez pour développer des outils thérapeutiques efficaces contre le SIDA.
[1] Barré-Sinoussi F., et al., Isolation of a T-Lymphotropic Retrovirus from a Patient at Risk for Acquired Immune Deficiency Syndrome (AIDS). Science, 1983. DOI : 10.1126/science.6189183 [Publication scientifique]
[2] Klatzmann D., et al., Selective tropism of lymphadenopathy associated virus (LAV) for helper-inducer T lymphocytes. Science, 1984. DOI : 10.1126/science.6328660 [Publication scientifique]
[3] Bensussan A., et al., Les kinoïdes: une nouvelle génération de vaccins thérapeutiques. Médecine/Sciences (Paris), 2008. DOI : 10.1051/medsci/2008243306 [Review]
[4] Bailes E., et al., The evolution of primate lentiviruses and the origins of AIDS. The Molecular epidemiology of human viruses, 2002. DOI : 10.1007/978-1-4615-1157-1_5 [Livre de science]
Publié le 28/09/2025
 Aurélien Schwob/Papier-Mâché/CC BY-NC-SA 4.0 2025
Aurélien Schwob/Papier-Mâché/CC BY-NC-SA 4.0 2025Texte et images.
Utilisation des corrélats immunitaires pour comprendre la progression du SIDA
Écriture : Aurélien Schwob
Relecture scientifique : Lise Chauveau et Paul Verlhac
Relecture de forme : Félicités Rapon et Eléonore Pérès
Temps de lecture : environ 20 minutes.
Thématiques : Immunologie et Microbiologie (Biologie)
Publication originale : Cumont MC., et al., Early divergence in lymphoid tissue apoptosis between pathogenic and nonpathogenic simian immunodeficiency virus infections of nonhuman primates. Journal of Virology, 2007. DOI : 10.1128/jvi.00450-07. Accès libre sur HAL (pdf).
Que ce soit pour résoudre un exercice complexe de géométrie, apprendre à pêcher ou à jouer à la belote, qui n’a jamais cherché de l’aide auprès d’une personne de sa famille ? Certains scientifiques ont poussé l’idée un peu plus loin pour tenter d’élucider certains mystères qui persistent autour de la pathologie du SIDA, en étudiant l’expérience de nos cousins les singes, qui ont un long vécu avec des virus proches. Loin d’une paisible partie de pêche, cette aventure pourrait bien remettre en cause ce que vous croyez savoir sur les tenants et aboutissants de cette maladie…
On attribue généralement à Confucius la maxime suivante : « L’homme sage apprend de ses erreurs, l’homme plus sage apprend des erreurs des autres ». Ce proverbe qui nous incite à profiter des expériences et enseignements de nos pairs, nous l’appliquons consciemment ou non toutes et tous au long de nos parcours de vie. C’est d’ailleurs une caractéristique humaine à l’échelle de l’évolution. En effet, notre espèce est notamment caractérisée par une longue phase juvénile au cours de laquelle les jeunes apprennent au contact de leur entourage. Plus précisément, cette caractéristique est commune à l’ensemble du groupe des primates. L’apprentissage joue un rôle primordial chez tous les singes, qu’ils soient humains ou non !
Au cours de cet article, nous allons voir comment prendre au mot cette maxime pour engranger des connaissances sur un thème particulièrement pointu : celui des agents pathogènes et des maladies infectieuses. Pour cela, nous allons nous pencher sur l’exploitation de corrélats immunitaires, un outil très puissant permettant d’élucider divers facteurs impliqués dans la progression pathologique des maladies infectieuses. Ainsi, en comparant un individu développant une pathologie suite à une infection à un autre individu ne développant pas de pathologie, il est possible d’identifier les facteurs immunitaires à l’origine de la sensibilité à ce pathogène. Mais cette approche semble restreinte aux cas d’infections aiguës identifiables précocement pour permettre le suivi des différents paramètres. Cela semble très difficile à appliquer au cas de l’infection par le Virus de l’Immunodéficience Humaine (VIH) et de la progression vers le Syndrome d’ImmunoDéficience Humaine (SIDA) qui s’ensuit. C’est dans ce contexte que plusieurs équipes de la communauté VIH ont entrepris depuis plusieurs années d’étudier les corrélats immunitaires entre hôtes humains infectés par le VIH et hôtes simiens infectés par leur Virus de l’Immunodéficience Simienne (VIS) respectifs. Une étude réalisée par une équipe française de l’INSERM en 2007 apporte, par une stratégie originale de corrélation immunitaire, un éclairage particulier sur cette question, notamment pour tenter de discriminer les effets liés au virus de ceux liés à l’hôte.
Étudier les facteurs de la progression vers le SIDA : le serpent qui se mord la queue
Le Syndrome d’ImmunoDéficience Acquise, ou SIDA est une maladie mortelle touchant l’espèce humaine. Elle est caractérisée par une phase d’immunodéficience, c’est-à-dire un état dans lequel notre système immunitaire n’est plus capable de défendre l’organisme contre les menaces externes (infections) ou internes (cancers). Lors de cette phase, les patients concernés développent des maladies dites opportunistes, c’est-à-dire des pathologies normalement très bien contrôlées par le système immunitaire et qui ne touchent donc que les immunodéprimés. Touchés par ces maladies opportunistes (comme divers cancers) et plus exposés aux autres pathogènes (virus ou autres bactéries), les patients décèdent précocement s’ils n’ont pas accès à des traitements adaptés.
L’origine de cette immunodéficience a été rapidement identifiée dès le début des études portant sur le SIDA, et notamment la découverte du lien entre cette maladie et l’infection par le Virus de l’Immunodéficience Humaine (VIH) par Françoise Barré-Sinoussi et Jean-Claude Chermann en 1982 [1]. Il s’est avéré que, suite à l’infection par le VIH, on observait une perte de fonction des lymphocytes T CD4 [2]. Cette catégorie de lymphocytes ayant comme fonction de coordonner les différents acteurs du système immunitaire adaptatif et de permettre leur activation, on comprend aisément pourquoi la chute de ce maillon de la chaîne engendre la déstructuration de l’ensemble de la réponse immunitaire (Figure 1). En effet, les lymphocytes T CD4 sont nécessaires à l’activation à la fois des lymphocytes T CD8 cytotoxiques, responsables de la destruction des cellules infectées, ainsi que des lymphocytes B producteurs d’anticorps.
Le problème auquel est confrontée la communauté des chercheurs sur ce domaine ne se situe pas tout-à-fait ici mais une étape en amont : comment l’infection par le VIH déclenche-t-elle cette perte de fonction des lymphocytes T CD4 ? Ces cellules étant des cibles de l’infection par le VIH, provoquant leur destruction, on pourrait se dire que le mécanisme est simple : le VIH va infecter un à un tous les lymphocytes T CD4 jusqu’à épuisement, amenant à leur destruction totale et donc à l’arrêt de l’immunité. Mais en réalité il n’est pas si clair que ce mécanisme soit suffisant pour expliquer l’épuisement des lymphocytes. En effet, au fil des ans, d’autres hypothèses ont émergé, comme une destruction indirecte de ces lymphocytes, une simple inactivation ou encore le résultat d’une réponse immunitaire trop élevée ou au contraire, trop faible (Figure 2).
Cette question est rendue particulièrement épineuse par différents paramètres. Tout d’abord, les cellules dont la fonction est de lutter contre le virus sont précisément ses cibles, ce qui rend complexe l’interprétation des changements les concernant. En effet, ces changements pourraient être des réactions permettant la lutte contre le pathogène ou au contraire un effet de l’infection. Mais le plus gros obstacle est le temps. En effet, le SIDA n’apparaît qu’après une période de progression lente de la pathogenèse se comptant le plus souvent en années, et le moment de l’infection initiale passe très souvent inaperçu. Pour contourner ces différents problèmes, les membres de la communauté scientifique qui s’attaquent à cette question exploitent un outil particulièrement puissant : les « corrélats immunologiques ». Comme nous l’avons déjà évoqué en introduction, il s’agit de l’identification de mécanismes immunitaires qui sont associés à la gravité de la pathologie telle qu’elle se développe réellement chez le patient et aident à l’expliquer. Pour illustrer avec un exemple un peu caricatural, considérons deux patients infectés par le même virus mais dont l’un survit et le second décède (Figure 3). Si après analyse de leur système immunitaire, on se rend compte que le patient ayant survécu a produit des anticorps spécifiques de ce virus et que le patient ayant trépassé n’a pas produit d’anticorps, on aura identifié un mécanisme immunitaire associé à la progression de la pathologie pour ce virus.
Cette étude étant rendue complexe chez l’hôte humain pour le cas du SIDA, plusieurs stratégies consistent à étudier l’infection des autres membres du groupe des primates auquel Homo sapiens appartient. Cette stratégie exploite le fait que nos cousins les singes possèdent des équivalents du VIH, que l’on appelle les VIS (qui pour rappel signifie Virus de l’Immunodéficience Simienne), et que les réactions des singes à l’infection par leur VIS respectif est parfois différente de celle des humains lors de l’infection par le VIH. Prenons par exemple l’espèce du Macaque Rhésus. Cette espèce vivant en Asie du Sud-Est peut être infectée par un VIS spécifique (le VIS macaque), ce qui provoque systématiquement une progression vers une phase SIDA et le décès des animaux infectés, exactement comme chez l’hôte humain. À l’inverse, chez le Singe Vert Africain, l’infection par son VIS spécifique (le VIS singe vert) ne provoque pas de pathologie. On a ainsi un modèle comparatif entre des infections pathologiques et non pathologiques, permettant de savoir quels sont les facteurs caractéristiques de la progression vers la pathologie.
Un oiseau rare pour lever le lièvre sur la maladie des singes…
Mais il reste tout de même un problème. En effet, il s’agit ici d’hôtes d’espèces différentes, infectés par des souches virales différentes. Comment exclure que les virus simiens étudiés n’aient pas simplement évolué d’une manière différente les uns par rapport aux autres ? Ces différences sont-elles révélatrices d’une réelle différence dans la manière dont l’hôte réagit à l’infection ou plus simplement de propriétés virales différentes ? Pire encore, il faut bien cultiver les virus dans des cellules pour obtenir les stocks de virus utilisés dans ces expériences. L’évolution virale étant très rapide, comment exclure l’hypothèse que ces virus se seraient évolutivement adaptés aux cellules utilisées lors de la production virale, ce qui expliquerait une réplication plus efficace ? Pour se tirer de ces deux ornières, l’équipe française que nous suivons dans ses aventures a décidé de modifier deux paramètres déterminants.
En premier lieu, ils ont intégré dans leur conception expérimentale deux populations de la même espèce : des Macaques Rhésus d’origine chinoise qui progressent lentement vers le SIDA après infection par leur VIS (la moitié des individus décède 35 mois après infection) et des Macaques Rhésus d’origine indienne qui progressent rapidement vers le SIDA (la moitié des individus est ici décimée en à peine 10 mois) (Figure 4).
Dans un second temps, l’équipe a testé l’hypothèse selon laquelle la réplication virale serait plus efficace dans la population indienne (dont le SIDA progresse plus rapidement que dans la population chinoise) parce que le virus leur serait adapté. Cette hypothèse repose sur le fait que, dans la plupart des études réalisées jusqu’à présent, les stocks de virus VISmacaque utilisés sont produits en cellules issues de Macaque Rhésus de la population indienne. Les scientifiques ont donc testé l’impact de la souche virale étudiée, en utilisant des virus VISmacaque produits pendant plusieurs cycles de réplication virale dans des cellules issues de Macaques Rhésus de la population chinoise. Si l’hypothèse est vraie, alors on s’attend à voir une inversion du résultat, c’est-à-dire une meilleure réplication chez les Macaques Rhésus chinois, sur les cellules desquelles nos virus ont été répliqués.
Des primates qui ne se contentent pas tous de singer l’infection humaine
Après avoir infecté des populations de Singe Vert Africain avec le virus VIS singe vert et une population de Macaques Rhésus de Chine ainsi qu’une population de Macaque Rhésus d’Inde toutes les deux avec la même souche de VIS macaque, l’équipe a mesuré un certain nombre de paramètres viraux et immunologiques sur ces animaux. Une fois les différences de progressions pathologiques exposées plus haut (Figure 4) vérifiées, la première chose à faire a été de mesurer la virémie, c’est-à-dire la quantité de virus circulant dans le sang. L’infection par les VIS, comme le VIH, se caractérise par une première phase dite aiguë où le virus se réplique beaucoup, suivie par une phase dite chronique où la virémie baisse. Les chercheurs ont donc mesuré principalement la virémie à son pic d’infection initiale. Cela permet également de chercher les événements précoces prédictifs d’une progression pathogénique plus ou moins rapide.
Dans la logique de l’hypothèse selon laquelle la destruction des lymphocytes est le résultat de la seule infection virale qui détruirait les cellules une à une, on s’attend à ce que la virémie soit proportionnelle à la pathologie : plus les virus infectent de cellules, plus ils se multiplient et plus les cellules immunitaires sont détruites (Figure 5).
Dans cette hypothèse, le SIDA serait provoqué par une réponse immunitaire trop faible pour contrer le virus. Or, lorsque l’équipe de recherche compare la virémie au pic de l’infection entre les trois conditions étudiées, ils observent que cette virémie n’est pas significativement différente entre elles ! En effet, que ce soit chez le Singe Vert Africain ne développant pas de pathologie, le Macaque Rhésus Chinois progressant lentement vers le SIDA ou le Macaque Rhésus Indien progressant rapidement vers le SIDA, on trouve toujours entre approximativement 1 million et 100 millions de virus par millilitre de sang ! Ces résultats, conformes à d’autres observations déjà réalisées à l’époque, portent un sérieux coup à l’hypothèse d’une destruction « manuelle » des cellules immunitaires par le virus… et rendent caduque l’idée de prédire la gravité de la progression pathologique en se basant sur la virémie seule.
Après cette première observation pouvant sembler étonnante, l’équipe de recherche a poussé plus loin ses investigations. Une hypothèse pour expliquer l’absence des lymphocytes T est que leur destruction se fasse par un processus d’apoptose. L’apoptose, ou « mort cellulaire programmée » est un phénomène biologique intégré au fonctionnement normal de l’organisme par lequel les cellules vont fragmenter leur ADN et se détruire « de l’intérieur ». Or, on sait que l’apoptose est un processus majeur de la physiologie immunitaire. Elle intervient notamment à la fin de la réponse immunitaire au niveau des lymphocytes T CD4 qui vont pour la plupart être détruits une fois leurs pathogènes-cibles éradiqués. Ce mécanisme constitue donc un bon candidat pour expliquer la destruction de cellules immunitaires.
Les scientifiques ont donc quantifié le phénomène d’apoptose à l’intérieur de ganglions lymphatiques, ces structures qui enflent lorsque nous sommes infectés et qui sont le lieu de rencontre, d’activation, et de multiplication des cellules immunitaires suite à la détection d’un pathogène. Par une technique baptisée TUNEL, permettant de colorer les cellules dont l’ADN est fragmenté, les chercheurs ont visualisé directement au microscope le nombre de cellules en apoptose dans les ganglions prélevés chez les différents animaux infectés. Les résultats montrent que l’apoptose augmente fortement au pic de l’infection chez les Macaques Rhésus Chinois, encore un peu plus chez les Macaques Rhésus Indiens alors que chez le Singe Vert Africain, l’apoptose n’augmente pas suite à l’infection. Confirmant des suppositions précédentes, l’équipe vient ainsi de mettre la main sur un facteur qui semble proportionnel à la progression pathogénique ! L’apoptose des lymphocytes au pic de l’infection (donc lors des phases précoces) est proportionnelle à la rapidité de la progression pathogénique. Si cela semble assez logique, il convient de noter un élément supplémentaire particulièrement riche d’enseignements une fois cette conclusion faite. Le fait que le taux d’apoptose soit décorrélé de la virémie montre bien que cette hausse de l’apoptose n’est pas directement liée au virus, et qu’il existe d’autres mécanismes au niveau cellulaire expliquant la progression vers la phase SIDA.
Quand le système immunitaire monte sur ses grands chevaux
Une fois le rôle de l’apoptose dans le processus de progression pathogène mis en évidence, il s’agit maintenant d’essayer de comprendre son origine. L’apoptose étant liée au niveau d’activation immunitaire, c’est l’un des paramètres qui a été exploré dans cette étude. Pour ce faire, les membres de l’équipe de recherche ont estimé le niveau d’activation des lymphocytes de nos trois conditions étudiées en se basant sur la détection des marqueurs qui sont situés à la surface des cellules lorsqu’elles sont activées. Ainsi, en marquant ces molécules de surface avec une molécule fluorescente, il est possible d’estimer le nombre de cellules activées par une technique de cytométrie en flux consistant pour faire simple à faire passer les cellules individuellement devant un lecteur de fluorescence automatisé. Cela permet de dénombrer les cellules portant les marqueurs de l’activation rendus fluorescents, et donc d’estimer le taux d’activation des cellules.
Lorsque l’on compare le niveau d’activation des lymphocytes entre nos trois types de singes, on observe tout d’abord que les deux Macaques Rhésus sont caractérisés par un niveau d’activation des lymphocytes bien plus élevés que le Singe Vert Africain (avec environ 4 fois plus de cellules actives au pic infectieux). Cela confirme que la faible apoptose des singes ne développant pas de pathologie SIDA est corrélée à un faible taux d’activation des lymphocytes par rapport aux singes développant une pathologie SIDA. De manière intéressante, on ne trouve pas de différence significative entre les deux populations de Macaque Rhésus, alors même qu’il y a une différence de progression pathologique. Avec ce résultat, on montre que cette approche de corrélats immunologiques chez les primates non-humains permet d’établir des corrélations entre les événements précoces de l’infection et la gravité de la progression pathologique.
Mais il ne s’agit pas de la seule différence observable entre Macaques Rhésus et Singes Verts Africains. En effet, les résultats de l’étude montrent également que les Macaques Rhésus ont un niveau d’activation de leurs lymphocytes cytotoxiques plus élevé, alors qu’au contraire les Singes Verts Africains présentent une plus grande activation des lymphocytes B, producteurs d’anticorps (Figure 6). Cela suggère qu’en plus de l’intensité de la réponse immunitaire, le type de réponse induite est également un paramètre important dans le contrôle de la pathogenèse.
Une cousinade qui permet de faire mouche
Les derniers résultats évoqués se concentrent principalement sur la comparaison entre Macaques Rhésus et Singes Verts Africains. Pourtant les chercheuses et chercheurs ayant réalisé l’étude en question ont pris la peine d’intégrer deux sous-populations de Macaques Rhésus, ainsi que d’utiliser un virus produit sur des cellules d’une de ces populations en particulier.
Si nous analysons les résultats obtenus en ayant ces éléments en tête, nous notons tout d’abord que, si le taux d’apoptose corrèle avec la progression vers le SIDA, les deux sous-populations de Macaques Rhésus ont pourtant le même niveau d’activation des lymphocytes (Figure 6). Cela suggère que l’apoptose n’est pas le seul paramètre lié à l’activation des lymphocytes T CD4 et que d’autres facteurs sont encore probablement à explorer.
Ce résultat suggère également qu’entre ces deux populations, c’est une différence au niveau de la manière dont leurs organismes réagissent à un même virus qui conditionne la pathologie. De plus, on constate que malgré l’utilisation de virus produits sur des cellules de Macaque Rhésus chinois pour infecter les deux populations simiennes, c’est bien la population indienne qui montre la progression la plus rapide. Cela ajoute encore un argument pour dire qu’il ne s’agit pas d’un effet lié à une variation de la souche virale qui est en jeu ici, mais plutôt à la réponse de l’hôte.
Conclusion : les singes qui murmurent à l’oreille des scientifiques
À l’aide de corrélats immunitaires, cette étude a donc approfondi nos connaissances dans la pathogenèse du VIH et sa progression jusqu’à la déclaration de la phase SIDA. En partant des données déjà disponibles sur les différences de vitesse de progression vers le SIDA de plusieurs espèces simiennes et sur l’idée déjà connue d’un potentiel lien entre cette progression et l’apoptose des lymphocytes T, les scientifiques à l’origine de cette étude ont permis d’élucider les mécanismes qui sous-tendent l’apoptose des lymphocytes T, permettant de faire progresser la question de l’origine de la phase SIDA lors d’une infection par un VIS.
En effet, en démontrant le lien entre apoptose des lymphocytes T et leur niveau d’activation, ces travaux ont participé à la démonstration que la pathogenèse du SIDA était liée, de manière contre-intuitive, à un excès de réponse immunitaire des organismes face au VIS. Les espèces qui ne développent pas de pathologie suite à l’infection ne sont pas particulièrement capables d’éradiquer le virus, mais sont caractérisées par une réponse immunitaire initiale moins forte. Pour simplifier, cette suractivation du système immunitaire va provoquer in fine son épuisement, là où une réponse plus modérée permet de sauvegarder les forces du système immunitaire. Et sans l’apport de nos cousins primates non-humains, difficile d’imaginer que le problème à l’origine d’une maladie caractérisée par une immunodéficience réside dans le fait d’avoir une réaction immunitaire trop forte !
Cette idée a depuis fait son chemin et fait maintenant consensus, à tel point que des pistes thérapeutiques sont envisagées consistant à modérer la réponse immunitaire des patients pour ralentir la progression SIDA [3].
Tout cela met en évidence la puissance de l’utilisation des corrélats immunitaires. Nous en avons ici un exemple utilisant des espèces non-humaines, mais de manière générale cette méthode a de nombreuses applications. Plutôt que d’essayer de comprendre les mécanismes sous-jacents dans un système d’interactions éminemment complexe, l’étude des corrélats se veut plus statistique. Prenons deux types de patients soumis au même agent pathogène, l’un développant une pathologie rapide et l’autre montrant peu de symptômes. En comparant leurs réponses immunitaires cela nous donnera autant de pistes de compréhension des mécanismes à l’œuvre, et même, des pistes thérapeutiques ! Ces approches sont d’ailleurs utilisées pour l’étude des infections humaines par le différents Virus de l’Immunodéficience Humaine, soit en comparant des patients contrôlant plus ou moins l’infection, soit en comparant les infections par les virus VIH-1 et VIH-2, aux pouvoirs pathogènes différents. Il reste que l’étude des primates non-humains reste un outil de choix pour l’étude des phases d’infection précoce, ainsi que pour les outils d’investigation plus invasifs permis par l’utilisation de modèles animaux.
Bien entendu, les conclusions de cette étude seule ne doivent pas nous emmener trop loin dans notre enthousiasme. Aussi brillante que soit l’idée de comparer nos cousins évolutifs infectés par les cousins évolutifs de nos virus, il faut garder en tête que des différences importantes peuvent exister entre ces espèces animales et virales. Ainsi, il serait possible que ces conclusions valables pour les interactions singes / VIS ne puissent s’appliquer au contexte de l’hôte humain / VIH. Cette réserve a donc nécessité des investigations complémentaires visant à confirmer ces hypothèses pour les hôtes humains. Mais la forte parenté entre primates non-humains et humains, ainsi que la parenté particulièrement forte entre VIS et VIH (le second dérivant récemment du premier [4]) permet d’accorder beaucoup de crédit à ces découvertes dans ce contexte précis.
Au final, en plus de nous apporter des réponses sur la pathogenèse des VIS et VIH, cette étude nous rappelle que lorsqu’il s’agit d’interactions hôtes/pathogènes, tout est question d’histoire évolutive commune et d’adaptation des organismes les uns aux autres. Cela montre encore une fois la place centrale des concepts évolutifs dans toutes les branches de la biologie. Reste à savoir si, comme Confucius nous y incite, ces découvertes nous permettront d’en apprendre assez pour développer des outils thérapeutiques efficaces contre le SIDA.
[1] Barré-Sinoussi F., et al., Isolation of a T-Lymphotropic Retrovirus from a Patient at Risk for Acquired Immune Deficiency Syndrome (AIDS). Science, 1983. DOI : 10.1126/science.6189183 [Publication scientifique]
[2] Klatzmann D., et al., Selective tropism of lymphadenopathy associated virus (LAV) for helper-inducer T lymphocytes. Science, 1984. DOI : 10.1126/science.6328660 [Publication scientifique]
[3] Bensussan A., et al., Les kinoïdes: une nouvelle génération de vaccins thérapeutiques. Médecine/Sciences (Paris), 2008. DOI : 10.1051/medsci/2008243306 [Review]
[4] Bailes E., et al., The evolution of primate lentiviruses and the origins of AIDS. The Molecular epidemiology of human viruses, 2002. DOI : 10.1007/978-1-4615-1157-1_5 [Livre de science]
Publié le 28/09/2025
Aurélien Schwob/Papier-Mâché/CC BY-NC-SA 4.0 2025Texte et images.