Chargement de l'article...
Écriture : Noémie Frezel-Jacob
Relecture scientifique : Alexandra Gros et Estelle Nakul
Relecture de forme : Pauline Bellot et Élodie Billard
Temps de lecture : environ 7 minutes.
Thématiques : Neurobiologie (Biologie)
Publication originale : Woo S.H., et al., Piezo2 is the principal mechanotransduction channel for proprioception. Nature neuroscience, 2015. DOI : 10.1038/nn.4162

En 2021, le prix Nobel de physiologie a récompensé la découverte de molécules permettant à nos neurones sensoriels de détecter les stimulations extérieures : le chaud, le froid, une pression mécanique, des éléments chimiques, mais aussi l’étirement ou la déformation des cellules. Grâce à l’une de ces molécules, appelée canal Piezo2, il nous est possible de tenir debout, de marcher ou de faire nos lacets. La publication scientifique présentée ici explique le rôle de ce canal.
La molécule de notre sixième sens
En grec, piezo signifie « pression ». Le canal Piezo2, découvert en 2010 [1] par l’équipe du Professeur Ardem Patapoutian, serait impliqué dans notre capacité à sentir les contraintes mécaniques (comme l’étirement ou la pression). Le fonctionnement de ce canal permettrait notamment de pouvoir situer nos membres dans l’espace. En 2015, les scientifiques auteurs de cette publication, de l’équipe d’Ardem Patapoutian, découvrent que cette même molécule est essentielle à notre « sixième sens » : la proprioception.
La proprioception est le sens qui nous permet de connaître la position de notre corps et de nos membres dans l’espace, et d’effectuer correctement la plupart de nos mouvements. Cela nous permet par exemple de marcher et de tenir debout même sur un sol inégal. Il existe deux types de proprioception :
- l’une dite inconsciente : qui comprend entre autres les réflexes ;
- et l’autre dite consciente : qui nous permet par exemple de toucher notre nez avec notre doigt ou de fermer une boutonnière les yeux fermés sans difficulté.
Ces deux types de proprioception sont traités différemment dans le cerveau [2], mais ont pour origine les mêmes neurones sensoriels, dit proprioceptifs. Tout comme les neurones du toucher, de la sensation de chaud ou de froid, ou les neurones de la douleur, ils représentent une classe de neurones particuliers [3].
Les neurones proprioceptifs innervent d’un côté les muscles, les tendons et les articulations, c’est-à-dire que les terminaisons sensorielles y sont connectées. Ces mêmes neurones se terminent de l’autre côté dans la moelle épinière et le tronc cérébral (Figure 1), d’où l’information perçue est envoyée jusqu’au cerveau.
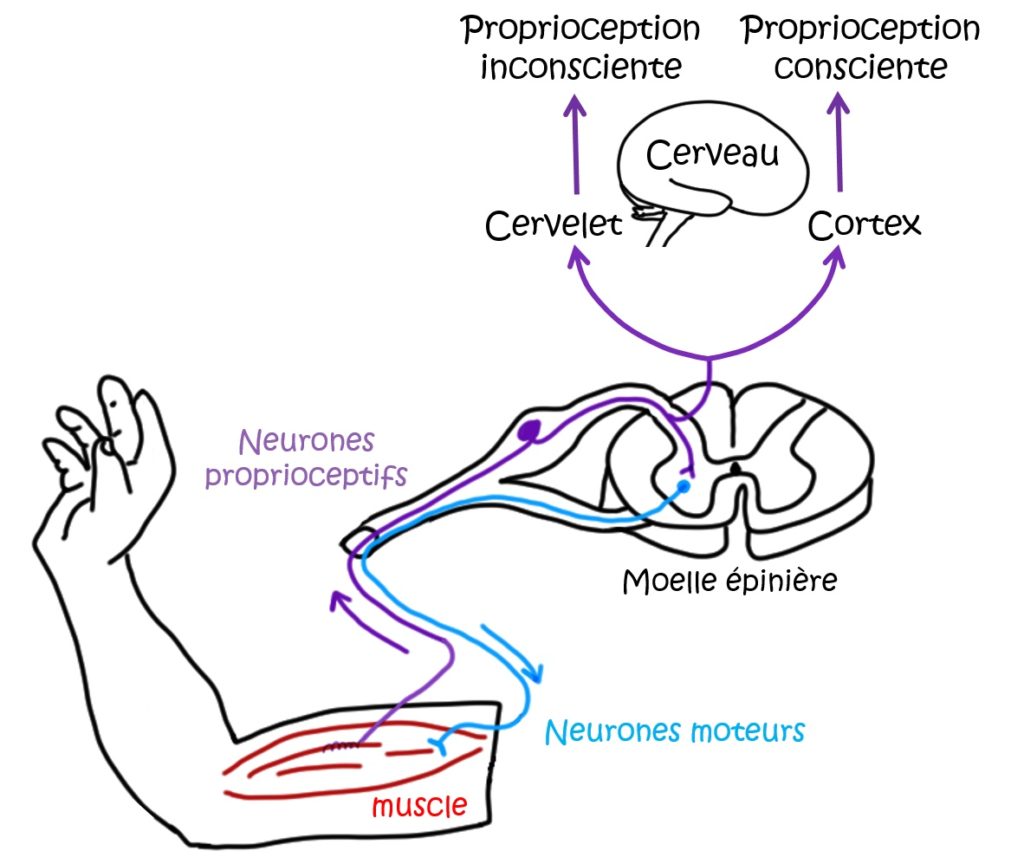
Comment nos muscles et articulations sont-ils capables de sentir leur position dans l’espace ?
Il s’agit en réalité de sentir les forces (ou déformations) mécaniques, telles que l’étirement, la torsion des muscles ou des tendons, ou bien le poids supporté par l’articulation. Tous ces paramètres sont détectés par les neurones sensoriels proprioceptifs, et l’information qui en résulte est analysée dans la moelle épinière et le cerveau. Pour détecter ces contraintes au niveau des muscles et des articulations, les neurones proprioceptifs doivent posséder un (ou plusieurs) récepteur(s) de pression. Et c’est exactement le rôle que joue le canal Piezo2.
À l’instar des pigments visuels, des récepteurs du goût et de l’odorat [4], connus depuis longtemps, il existe aussi des récepteurs pour le chaud, le froid ou encore pour la douleur. Avant 2010, on ne connaissait pas les récepteurs pour le toucher et la proprioception. La découverte des canaux de la famille Piezo (Piezo1 et Piezo2) a alors changé la donne [1]. On connaît à présent des molécules (ou récepteurs) capables de détecter des contraintes mécaniques appliquées à la surface des cellules. Mais ces récepteurs sont-ils également capables de détecter des déformations comme l’étirement au niveau des muscles ?
Piezo2 permet aux neurones proprioceptifs de sentir l’étirement des muscles
En utilisant diverses méthodes d’enregistrement, de traçage et de manipulation des neurones sensoriels, les chercheur·ses démontrent que Piezo2 est présent dans les neurones proprioceptifs des muscles, ce qui les rend « sensibles » à la pression ou à l’étirement. En fait, quand un muscle est étiré (comme l’est notre biceps lorsque l’on tend le bras), les neurones situés à l’intérieur du muscle se retrouvent également étirés. Piezo2, qui est localisé sur ces neurones, permet de détecter l’étirement et d’activer les neurones. Ces derniers envoient l’information « le biceps est étiré, le bras doit être tendu » jusqu’à la moelle épinière (ou le tronc cérébral [*]) et au cerveau.
Ce type de stimuli est détecté dans les muscles au niveau de structures spécialisées qui contiennent les terminaisons des neurones proprioceptifs. Pour les membres postérieurs, les parties des neurones qui transmettent les messages sont regroupées dans le nerf sciatique (Figure 2). Si l’on enregistre l’activité électrique au niveau de ce nerf, on observe une augmentation des « réponses » lorsque l’on étire le muscle correspondant. La même expérience répétée avec des muscles provenant de souris chez qui le canal Piezo2 a été supprimé, ne produit presque plus de réponses (Figure 2, en rouge). La présence de Piezo2 dans les neurones proprioceptifs des muscles est donc bien nécessaire pour détecter l’étirement et générer le message qui sera envoyé au reste du système nerveux.
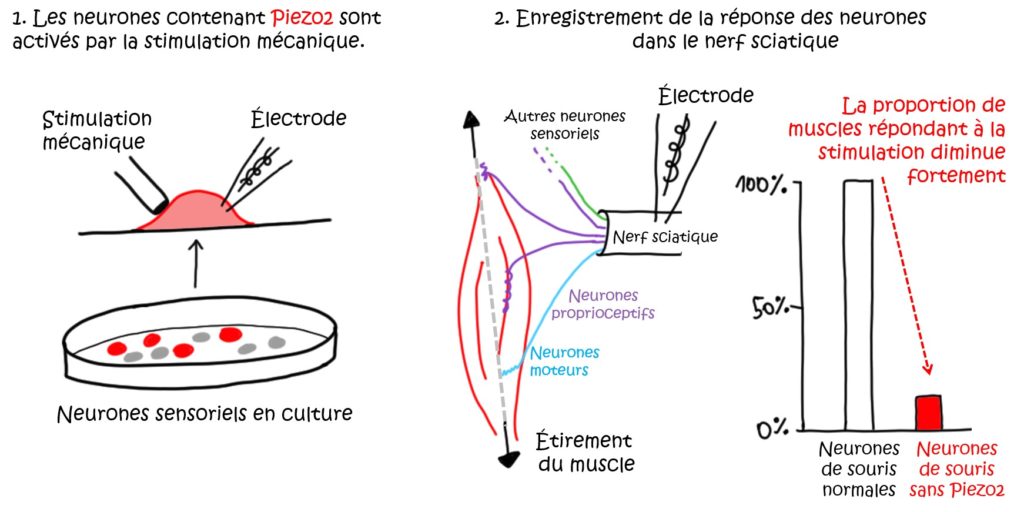
Les souris qui n’ont pas Piezo2 ont aussi des problèmes d’équilibre
Les résultats précédents montrent que sans Piezo2, l’étirement des muscles et des tendons n’est pas détecté par le système nerveux. Or la moelle épinière (ou le tronc cérébral [*]) a besoin de ces informations pour commander au reste des muscles les ajustements nécessaires pour la bonne réalisation du mouvement. Le cerveau utilise également ces informations pour déterminer la position de nos membres (c’est la proprioception consciente).
On peut donc faire l’hypothèse que des souris sans Piezo2 devraient présenter un certain nombre de déficits sensoriels et moteurs. C’est effectivement le cas. En effet, lorsque le canal Piezo2 est supprimé des neurones proprioceptifs, bien que le nombre et l’aspect général de ces neurones soient globalement normaux, les souris présentent de sévères déficits dès l’âge d’une semaine : leur démarche est mal coordonnée et la position de leurs membres est anormale.
Conclusion : un canal aux multiples fonctions et un prix Nobel
Les autrices et auteurs de cette étude montrent que le canal Piezo2 joue un rôle majeur dans la proprioception chez les souris, après avoir démontré quelques années auparavant qu’il est également nécessaire au sens du toucher [5].
L’identification de Piezo2 et de son rôle dans diverses fonctions biologiques liées, entre autres, à nos sens du toucher et de l’équilibre, est fondamentale à notre compréhension du corps humain et de nombreuses maladies. Ces découvertes ont ouvert la voie à de très nombreuses autres études sur Piezo1 et Piezo2 chez les souris mais aussi chez les humains. En particulier, il a été découvert que des patients souffrant de déficits profonds de la proprioception, de perturbation de l’équilibre et de la coordination motrice (ataxie), ou de mouvements asymétriques, avaient une anomalie au niveau des canaux Piezo2 [6]. Les molécules de la famille Piezo sont aussi présentes dans de nombreux autres organes. Piezo1 a notamment été identifié dans les globules rouges, les poumons, la vessie, le pancréas, et joue un rôle dans le fonctionnement du système vasculaire (Figure 3). Piezo2, en plus de la peau, des muscles et des tendons (pour son rôle dans le toucher et la proprioception) est aussi présent dans les poumons et le tube digestif [7] (Figure 3).
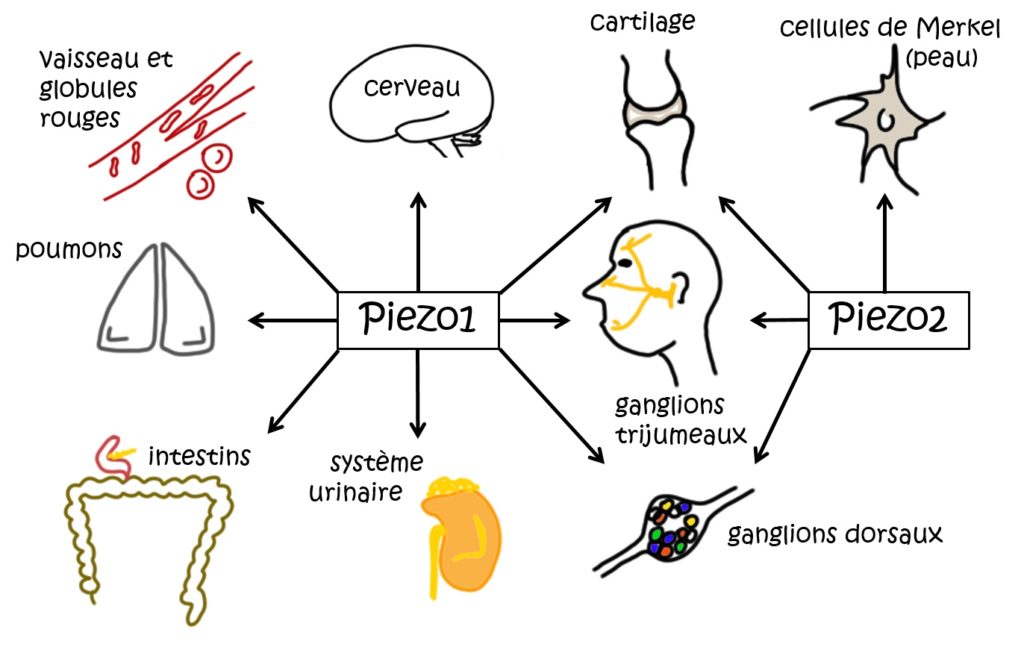
Ces travaux ont permis, après ceux, pionniers, de l’équipe de David Julius [8], d’expliquer les bases moléculaires de notre capacité à sentir notre environnement, et fournissent déjà des pistes dans le traitement de certaines maladies. C’est pourquoi ils ont permis à ces deux chercheurs, David Julius et Ardem Patapoutian, d’obtenir conjointement le prix Nobel de physiologie en 2021 [8].
[*] De manière générale, les neurones innervant les membres inférieurs et le tronc terminent dans la moelle épinière, et ceux innervant les membres antérieurs et le haut du corps plutôt dans le tronc cérébral.
[1] Coste B., et al., Piezo1 and Piezo2 Are Essential Components of Distinct Mechanically Activated Cation Channels. Science, 2010. DOI : 10.1126/science.1193270. [Publication]
[2] Delhaye B.P., et al., Neural Basis of Touch and Proprioception in Primate Cortex. Comparative Physiology, 2018. DOI : 10.1002/cphy.c170033. [Review]
[3] Lallemend F. & Ernfors P., Molecular interactions underlying the specification of sensory neurons. Trends in Neurosciences, 2012. DOI : 10.1016/j.tins.2012.03.006. [Review]
[4] Wicher D., et al., Design principles of sensory receptors. Frontiers in Cellular Neurosciences, 2010. DOI : 10.3389/fncel.2010.00025. [Article éditorial]
[5] Ranade S., Woo S.H., Dubin A., et al., Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature, 2014. DOI : 10.1038/nature13980. [Publication]
[6] Chesler A.T., et al., The Role of PIEZO2 in Human Mechanosensation. New England Journal of Medicine, 2016. DOI : 10.1056/NEJMoa1602812. [Publication]
[7] Fang G.Z., et al., Structure, kinetic properties and biological function of mechanosensitive Piezo channels. Cell and Biosciences, 2021. DOI : 10.1186/s13578-020-00522-z. [Review]
[8] The Nobel Prize in Physiology or Medicine 2021. Nobel Prize Outreach AB, 2022. [Article de presse, en anglais]
Publié le 12/03/2023
 Noémie Frezel-Jacob/Papier-Mâché/CC BY-NC-SA 4.0 2023
Noémie Frezel-Jacob/Papier-Mâché/CC BY-NC-SA 4.0 2023Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.
Écriture : Noémie Frezel-Jacob
Relecture scientifique : Alexandra Gros et Estelle Nakul
Relecture de forme : Pauline Bellot et Élodie Billard
Temps de lecture : environ 12 minutes.
Thématiques : Neurobiologie (Biologie)
Publication originale : Woo S.H., et al., Piezo2 is the principal mechanotransduction channel for proprioception. Nature neuroscience, 2015. DOI : 10.1038/nn.4162
En 2021, le prix Nobel de physiologie a récompensé la découverte de molécules permettant à nos neurones sensoriels de détecter les stimulations extérieures : le chaud, le froid, une pression mécanique, des éléments chimiques, mais aussi l’étirement ou la déformation des cellules. Grâce à l’une de ces molécules, appelée canal Piezo2, il nous est possible de tenir debout, de marcher ou de faire nos lacets. La publication scientifique présentée ici explique le rôle de ce canal.
Un nouveau rôle pour le canal ionique Piezo2
Cette étude datant de 2015 fait partie des travaux qui ont valu au professeur Ardem Patapoutian de partager avec David Julius le prix Nobel de physiologie et de médecine en 2021 [1]. Les scientifiques de son équipe ont montré que le récepteur Piezo2 est la molécule qui permet aux neurones sensoriels de détecter la déformation ou l’étirement des muscles. Cette équipe a également réussi à identifier ce canal ionique en 2010 [2], en montrant que les canaux ioniques Piezo1 et Piezo2 (piezo signifiant « pression » en grec) permettent à des neurones sensoriels en culture de sentir des stimulations mécaniques. Quelques années plus tard, ils démontrent que Piezo2 en particulier est essentiel au sens du toucher chez les souris [3]. Puis, en 2015, ils découvrent que cette même molécule est aussi essentielle à notre « sixième sens » qu’est la proprioception. Sans lui, les souris étudiées ont des mouvements mal coordonnés et des postures anormales, que l’on peut qualifier de déficits proprioceptifs.
La proprioception est le sens qui nous permet de connaître la position de notre corps et de nos membres dans l’espace, et d’effectuer correctement la plupart de nos mouvements. Cela nous permet par exemple de marcher et de tenir debout même sur un sol inégal. Il existe une proprioception dite inconsciente, qui comprend entre autres les réflexes (myotatique, posturaux, etc.), et une dite consciente, qui nous permet par exemple de toucher notre nez avec le doigt ou de fermer une boutonnière les yeux fermés et sans difficulté. Ces deux types de proprioception sont traités différemment dans le cerveau [4], mais ont pour origine les mêmes neurones sensoriels, dit proprioceptifs en périphérie. Tout comme les neurones du toucher, de la thermosensation (température), ou les neurones nociceptifs (douleur), ils représentent une classe de neurones particuliers [5]. Leurs corps cellulaires se situent dans les ganglions dorsaux, et ils innervent d’une part la périphérie (au niveau des muscles, tendons et des articulations) et se terminent d’autre part dans la moelle épinière et le tronc cérébral (Figure 1).
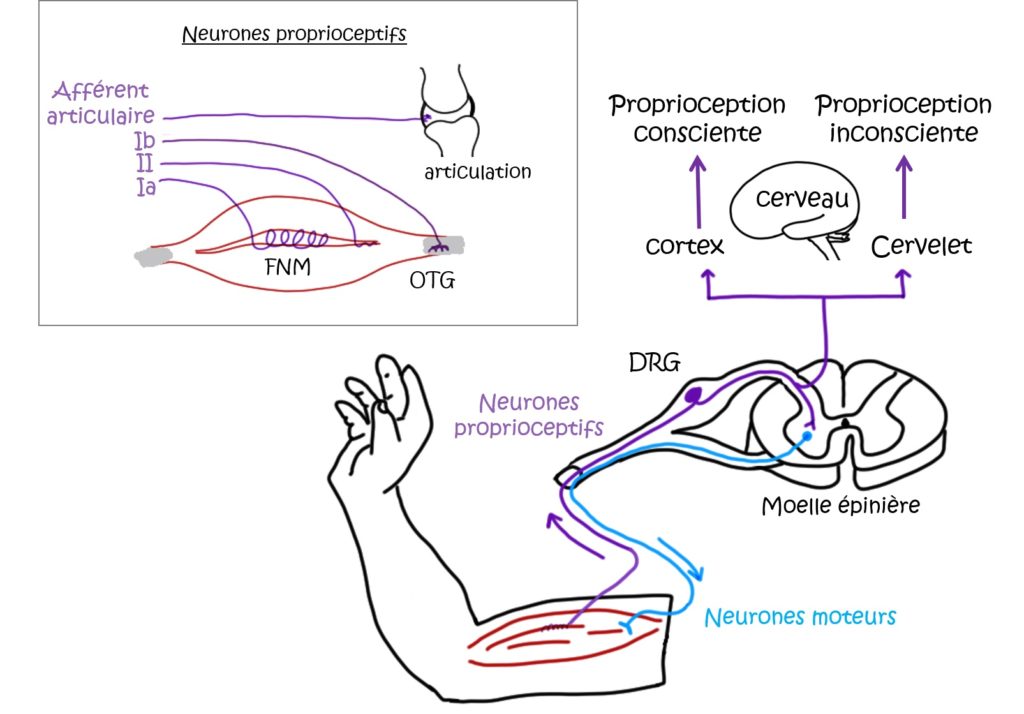
Comment nos muscles et nos articulations sont-ils capables de sentir leur position dans l’espace ?
Il s’agit en réalité de sentir des contraintes (ou forces) mécaniques, telles que l’étirement, la torsion des fibres musculaires ou des tendons, le poids supporté par l’articulation. Tous ces paramètres dits cinétiques sont détectés par les neurones proprioceptifs, et l’information qui en résulte est analysée dans la moelle épinière et le cerveau. Pour détecter ces paramètres en périphérie, c’est-à-dire au niveau des muscles et des articulations, les neurones proprioceptifs doivent posséder un (ou plusieurs) récepteur(s) de force. Et c’est exactement le rôle que joue la protéine Piezo2. On connaît depuis longtemps les pigments visuels ainsi que les récepteurs du goût et de l’odorat [6]. Depuis le début des années 1990 et l’identification des canaux de la famille TRP (pour Transient Receptor Potential) [7 ; 8], de très nombreux travaux ont permis d’identifier les neurones et les molécules nécessaires à la thermosensation (sentir le froid et le chaud), à l’audition, au toucher, à la douleur, etc. Sur la base de ces résultats, l’identification de la (ou des) molécule(s) responsable(s) de la sensation des contraintes mécaniques se concentraient sur deux autres familles de canaux [8]. Mais la découverte des canaux de la famille Piezo (Piezo1 et Piezo2) en 2010 [2] a changé la donne. On connaît à présent des canaux ioniques capables de détecter des contraintes mécaniques appliquées à la surface des cellules. Ces récepteurs sont-ils également capables de détecter les contraintes mécaniques comme l’étirement au niveau des muscles ?
L’utilisation de plusieurs lignées de souris transgéniques permet de démontrer le rôle du canal Piezo2 dans les neurones sensoriels proprioceptifs
Dans cette étude, plusieurs lignées de souris transgéniques ont été utilisées :
| Nom de la lignée | Caractéristiques |
| Pvalb-Cre;AI9tdtomato | Les neurones proprioceptifs expriment la protéine fluorescente rouge tdTomato (pour plus détails, voir recombinase et rapporteure) et vont donc être visibles en rouge. |
| Piezo2GFP | Le canal Piezo2 est fusionné à la protéine fluorescente verte, la GFP (on observe donc du vert aux endroits où le canal Piezo2 est présent). |
| Pvalb-Cre;Piezo2cKO [*] | Le canal Piezo2 est absent uniquement des neurones proprioceptifs. |
| Hoxb8-Cre;Piezo2cKO [*] | Le canal Piezo2 est absent d’une large population de neurones sensoriels situés dans la partie caudale de la colonne vertébrale (c’est-à-dire innervant le bas du dos et les pattes arrière). |
Pour mettre en évidence le rôle de Piezo2 dans la proprioception, Seung-Hyun Woo et ses collègues ont d’abord cherché à savoir si cette protéine est exprimée et fonctionnelle dans les neurones proprioceptifs. Pour cela, la première étape a été d’identifier ces neurones.
Les neurones proprioceptifs expriment une autre protéine appelée Parvalbumine (notée Pvalb ; on désigne comme « neurones Pvalb+ » les neurones qui l’expriment). Les scientifiques ont donc utilisé ce que l’on appelle une lignée de souris « rapporteure » : Pvalb-Cre;AI9tdtomato (Tableau 1). Dans ces souris, les neurones proprioceptifs (Pvalb+) apparaissent en rouge, et sont donc plus faciles à identifier et à étudier. Après avoir extrait et mis en culture ces neurones, ils ont pu enregistrer in vitro leur activité électrique (c’est-à-dire analyser s’ils sont activés et comment ils réagissent ; voir électrophysiologie) lors d’une stimulation mécanique du corps cellulaire (Figure 2, A). La très grande majorité des neurones (92 %) répondent à cette stimulation par un courant ionique appelé courant RA (pour Rapidely Adapting) (Figure 2, B). Lors de ces courants, le neurone est dépolarisé par la stimulation (des ions entrent dans la cellule, provoquant une inversion locale du potentiel de membrane), puis son potentiel de membrane revient très rapidement à sa valeur de base [**].
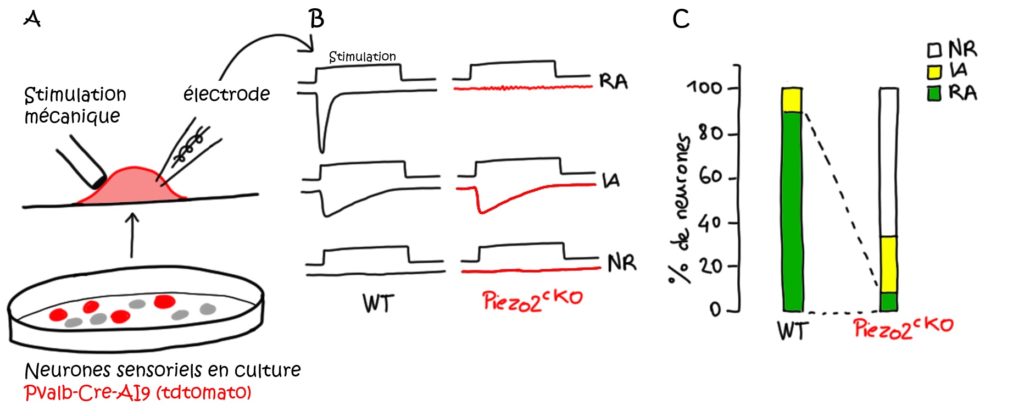
Ces courants RA sont composés principalement d’ions sodium Na+. Or, ces caractéristiques ressemblent de près à celles du canal ionique Piezo2 identifié quelques années auparavant [2]. En utilisant alors les souris Piezo2GFP (Tableau 1) les chercheurs montrent que Piezo2 est effectivement présent au niveau du corps cellulaire, mais aussi des terminaisons périphériques des neurones proprioceptifs Pvalb+, dans les fuseaux neuromusculaires et les organes tendineux de golgi (c’est-à-dire là où s’appliquent les contraintes mécaniques en conditions réelles). Pour cela, ils ont disséqué les muscles et les tendons de ces souris et révélé la présence des neurones Pvalb+ par immunofluorescence. Puis, ils ont observé que la fluorescence verte (GFP), indiquant la présence de Piezo2, est située au niveau des neurones fluorescents pour Pvalb, donc les neurones proprioceptifs. Mais la présence de Piezo2 dans ces neurones ne suffit pas à prouver que cette protéine y est nécessaire, ou ce qu’elle y fait [***]. Les expériences suivantes ont donc eu pour but de déterminer si Piezo2 confère la mécanosensibilité aux neurones proprioceptifs.
Piezo2 est responsable des courants mécanosensibles dans les neurones Pvalb+.
Les chercheurs ont alors enregistré l’activité électrique in vitro de neurones isolés soit de souris contrôles (WT), soit de souris Pvalb-Cre;Piezo2cKO, c’est-à-dire des souris chez qui Piezo2 a été supprimé sélectivement des neurones Pvalb+ (Tableau 1). Dans les neurones où Piezo2 est absent, les courants mécanosensibles RA sont très fortement diminués (seulement 8 % y sont encore présents), et 80 % des neurones ne répondent plus à une stimulation mécanique (Figure 2, C). Ceci démontre que Piezo2 est nécessaire à la transduction mécano-électrique, c’est-à-dire pour que ces neurones soient capables de générer une activité électrique en réponse à une stimulation mécanique.
Piezo2 est nécessaire pour détecter l’étirement des fibres musculaires innervées par les neurones Pvalb+
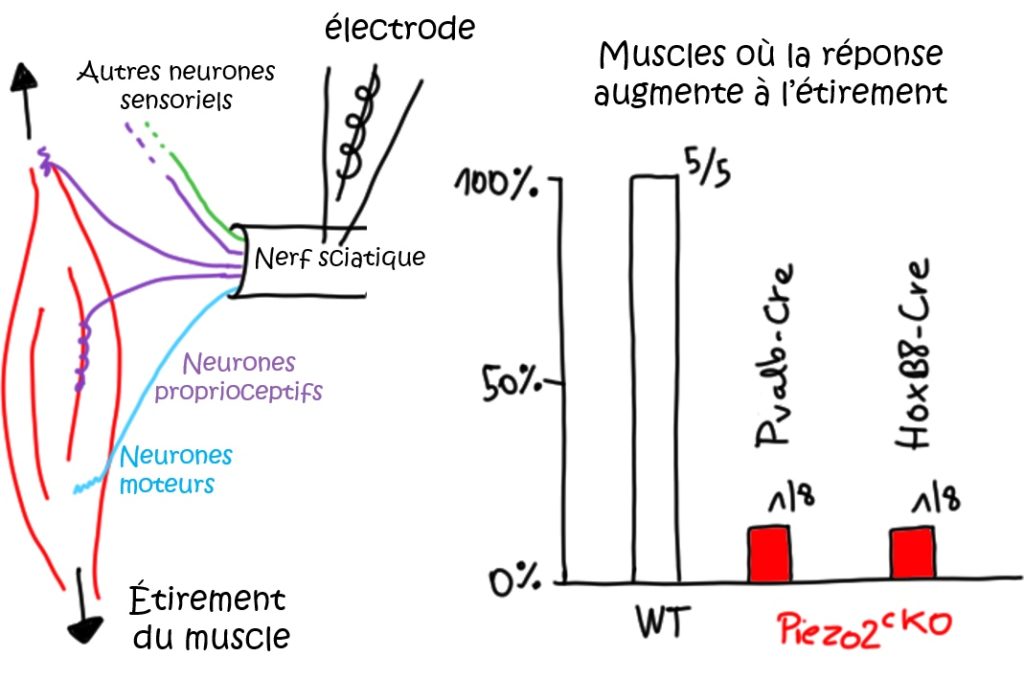
Néanmoins, ce résultat a été obtenu après stimulation directe du corps cellulaire des neurones proprioceptifs en condition in vitro. Or, chez l’animal, ces stimuli sont détectés en périphérie. Il existe deux principaux types d’organes mécanorécepteurs : les fuseaux neuromusculaires (FNM) et les organes tendineux de Golgi au niveau des tendons (OTG) (Figure 1). Ces organes sont innervés par la branche périphérique des neurones proprioceptifs. Au niveau des pattes arrières des souris (ou des jambes chez les humains), leurs axones sont regroupés avec les axones périphériques d’autres neurones sensoriels et des neurones moteurs, formant le nerf sciatique (Figure 3). En utilisant une préparation « nerf-muscle », les chercheur·ses ont enregistré l’activité électrique des neurones du nerf sciatique innervant un muscle de la patte arrière (Figure 3). Ils identifient d’abord les zones où se situent les réponses à l’étirement du muscle dans deux lignées de souris : dans la lignée Pvalb-Cre;Piezo2cKO, Piezo2 est supprimé des neurones proprioceptifs Pvalb+ ; dans la lignée HoxB8-Cre;Piezo2cKO, Piezo2 est supprimé d’une large population de neurones sensoriels situés dans la partie caudale de la colonne vertébrale (innervant le bas du dos et les pattes arrières) (Tableau 1). En l’absence de Piezo2 (muscles issus de souris de ces deux lignées), ils montrent que l’activité de ces neurones est fortement réduite, et qu’ils ne sont plus capables de détecter l’étirement des fibres musculaires. Piezo2 est donc également nécessaire à la transduction mécano-électrique en périphérie.
Piezo2 est aussi nécessaire pour la proprioception des souris in vivo
Les résultats précédents montrent que la présence du canal ionique Piezo2 dans les neurones proprioceptifs est nécessaire pour les rendre mécanosensibles in vitro. On peut donc faire l’hypothèse que des souris sans Piezo2 devraient présenter un certain nombre de déficits en termes de mécanosensibilité. Pour répondre à cette question, les chercheur·ses ont étudié les deux lignées de souris déjà citées (Pvalb-Cre;Piezo2cKO et HoxB8-Cre;Piezo2cKO), permettant de cibler les neurones proprioceptifs dès le début du développement [****]. Dans les deux cas, bien que le nombre et la morphologie des neurones proprioceptifs soient globalement normaux, les souris sans Piezo2 présentent de sévères déficits dès l’âge d’une semaine : leur démarche est mal coordonnée et la position de leurs membres est anormale. Ces observations sont semblables à celles effectuées chez des souris dont les neurones proprioceptifs ont été détruits [9].
Conclusion : un canal aux multiples fonctions et un prix Nobel
Les autrices et auteurs de cette étude montrent que le canal Piezo2 joue un rôle majeur dans la proprioception chez les souris, après avoir démontré qu’il est également nécessaire au sens du toucher [3]. Même s’ils n’excluent pas totalement l’existence d’un autre canal mécanosensitif, leurs résultats démontrent que Piezo2 est indispensable dans les neurones proprioceptifs, en particulier ceux innervant les fuseaux neuromusculaires. Il existe d’autres neurones proprioceptifs innervant les tendons et les articulations. Même si ceux-ci n’ont pas été étudiés spécifiquement comme l’ont été ceux des fuseaux neuromusculaires, il est très probable que Piezo2 y joue un rôle similaire. L’identification de Piezo2 et de son rôle dans diverses fonctions biologiques liées à la mécanosensation est fondamentale à notre compréhension du corps humain et de nombreuses maladies. Ces découvertes ont ouvert la voie à de très nombreuses autres études sur Piezo1 et Piezo2 chez les souris mais aussi chez les humains. En particulier, il a été découvert que des patients souffrant de déficits profonds de la proprioception, d’ataxie (perturbation de l’équilibre et de la coordination motrice) et de mouvements asymétriques avaient une mutation dans le gène codant pour Piezo2 [10]. Les canaux Piezo sont aussi présents dans de nombreux autres organes. Piezo1 a notamment été identifié dans les globules rouges, les poumons, la vessie, le pancréas, et joue un rôle dans le fonctionnement du système vasculaire (Figure 4). Piezo2, en plus de la peau, des muscles et des tendons (pour son rôle dans le toucher et la proprioception) est aussi présent dans les poumons et le tube digestif [11] (Figure 4).
Ces travaux ont permis, après ceux, pionniers, de l’équipe de David Julius, d’expliquer les bases moléculaires de notre capacité à sentir notre environnement. C’est pourquoi ils ont permis à ces deux chercheurs, David Julius et Ardem Patapoutian, d’obtenir conjointement le prix Nobel de physiologie 2021 [1].
[*] Les deux dernières lignées sont appelées KO conditionnel (ou cKO) car la suppression de Piezo2 dépend de la présence de la recombinase Cre. Dans les souris Pvalb-Cre, Cre est présente uniquement dans les neurones exprimant naturellement le gène Pvalb, et dans les souris Hoxb8-Cre, Cre est présente dans les neurones exprimant naturellement HoxB8. Piezo2 sera donc éliminé sélectivement de l’une ou l’autre de ces populations de neurones.
[**] Les propriétés électriques (le potentiel de membrane au repos, le type de réponse à une stimulation, etc.) sont l’un des nombreux moyens utilisés par les neuroscientifiques pour classifier les neurones en différents types (ou sous-populations). D’autres critères incluent la morphologie ou l’expression de certains gènes marqueurs. Ces critères peuvent renseigner sur la fonction des neurones concernés mais sont souvent utilisés principalement à des fins de classification.
[***] Afin d’identifier ou démontrer le rôle d’une structure (cela peut être une molécule, un type cellulaire, un tissu, un organe, etc.), la procédure classique est de supprimer cette structure. Si la fonction disparaît ou est altérée, c’est la preuve que la structure était nécessaire pour cette fonction en particulier : si Piezo2 est absent et que les neurones sont incapables de détecter une stimulation mécanique, alors Piezo2 est nécessaire à la détection de ce type de stimulation. Des expériences contrôles adaptées dans chaque cas sont bien sûr nécessaires.
[****] En 2014, l’équipe du professeur Patapoutian démontre qu’en éliminant Piezo2 d’une large population de neurones sensoriels chez la souris adulte (87 %, [2]), on observe des déficits tactiles et moteurs chez les souris. Afin d’étudier plus particulièrement l’impact de la suppression de ce canal pour la proprioception, les scientifiques ciblent à présent seulement les neurones proprioceptifs grâce à ces lignées de souris Pvalb+. À la différence de l’étude précédente, Piezo2 est ici supprimé des neurones ciblés dès la naissance, ce qui produit probablement un effet plus important que dans l’étude précédente.
[1] The Nobel Prize in Physiology or Medicine 2021. Nobel Prize Outreach AB, 2022. [Article de presse, en anglais]
[2] Coste B., et al., Piezo1 and Piezo2 Are Essential Components of Distinct Mechanically Activated Cation Channels. Science, 2010. DO : 10.1126/science.1193270. [Publication]
[3] Ranade S., Woo S.H., Dubin A., et al., Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature, 2014. DOI : 10.1038/nature13980. [Publication]
[4] Delhaye B.P., et al., Neural Basis of Touch and Proprioception in Primate Cortex. Comparative Physiology, 2018. DOI : 10.1002/cphy.c170033. [Review]
[5] Lallemend F. & Ernfors P.,Molecular interactions underlying the specification of sensory neurons. Trends in Neurosciences, 2012. DOI : 10.1016/j.tins.2012.03.006. [Review]
[6] Wicher D., et al., Design principles of sensory receptors. Frontiers in Cellular Neurosciences, 2010. DOI : 10.3389/fncel.2010.00025. [Article éditorial]
[7] Caterina M.J., et al., The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature, 1997. DOI : 10.1038/39807. [Publication]
[8] Venkatachalam K., et al., TRP channels. Annual Review of Biochemistry, 2007. DOI : 10.1146/annurev.biochem.75.103004.142819. [Review]
[9] Bewick G.S. & Banks R.W., Mechanotransduction in the muscle spindle. Pflugers Arch., 2015. DOI : 10.1007/s00424-014-1536-9. [Review]
[10] Chesler A.T., et al., The Role of PIEZO2 in Human Mechanosensation. New England Journal of Medicine, 2016. DOI : 10.1056/NEJMoa1602812. [Publication]
[11] Fang G.Z., et al.,Structure, kinetic properties and biological function of mechanosensitive Piezo channels. Cell and Biosciences, 2021. DOI : 10.1186/s13578-020-00522-z. [Review]
Publié le 12/03/2023
Noémie Frezel-Jacob/Papier-Mâché/CC BY-NC-SA 4.0 2023Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.