Chargement de l'article...
Ou comment se développe la forme des plantes
Écriture : Hervé Rabillé
Relecture scientifique : Cécile Feuillie et Éléa Hébérlé
Relecture de forme : Charlotte Rémy et Eléonore Pérès
Temps de lecture : environ 14 minutes.
Thématiques : Développement, Biologie végétale, Biologie cellulaire (Biologie)
Publication originale : Uyttewaal M., et al., Mechanical stress acts via katanin to amplify differences in growth rate between adjacent cells in Arabidopsis. Cell, 2012. DOI : 10.1016/j.cell.2012.02.048. Accès libre sur HAL (pdf).

Au sommet des tiges des plantes, de nouveaux organes aériens aux formes précises apparaissent en continu. Mais la croissance d’un grand nombre de cellules végétales s’accompagne d’irrémédiables conflits mécaniques, car elles sont entourées d’une solide paroi qui les lient les unes aux autres. En 2012, des chercheur·e·s ont montré, dans la petite plante Arabidopsis thaliana, que le stress mécanique accentue les écarts de croissance entre cellules voisines au lieu de les atténuer. Plus surprenant encore, cela semble faciliter le développement des formes de la plante !
Notre vie quotidienne est toujours plus ou moins stressante. Quel que soit le style de vie que nous menons, il y a toujours des besoins vitaux que nous devons satisfaire, pour nous et nos proches, avec des risques et des aléas, le futur n’étant jamais totalement prévisible. Voilà qui génère chez nous du stress psychologique. Nos manières personnelles de répondre à ce stress (et aux aléas de la vie, d’une manière générale) se traduisent par des différences dans le parcours de vie que nous suivons, tout au long de notre existence. En somme, la société est un entremêlement hétérogène de destins différents.
Or, dans le monde vivant, il n’y a pas qu’à l’échelle des populations (humaines, animales, végétales, etc.) qu’une telle hétérogénéité se manifeste. Chaque animal ou plante est, en quelque sorte, une mini société composée d’un très grand nombre de cellules aux fonctions différentes — ce sont des organismes dit pluricellulaires. Les cellules sont les unités de base de tous les êtres vivants sur Terre. Le plus souvent microscopique, la cellule est constituée a minima d’un milieu visqueux — le cytoplasme, qui contient, entre autres, le matériel génétique — entouré d’une membrane lipidique, qu’on appelle membrane plasmique, permettant de séparer la cellule du monde extérieur et de garantir son intégrité.
Les cellules végétales, aussi, ont la pression
Prenons les plantes, par exemple. De l’herbe la plus humble jusqu’au plus majestueux des chênes, elles sont toutes constituées de tissus, eux-mêmes composés de milliards de cellules. Au cours du développement, les cellules se reproduisent par division (une cellule-mère donnant deux cellules-filles) puis grandissent en augmentant en volume. Les tissus, comme les sociétés humaines, s’agrandissent par la multiplication et la croissance d’unités individuelles qui assurent, par leur comportement collectif, la mise en forme des organes : tiges, racines, feuilles, fleurs, fruits, etc. [1].
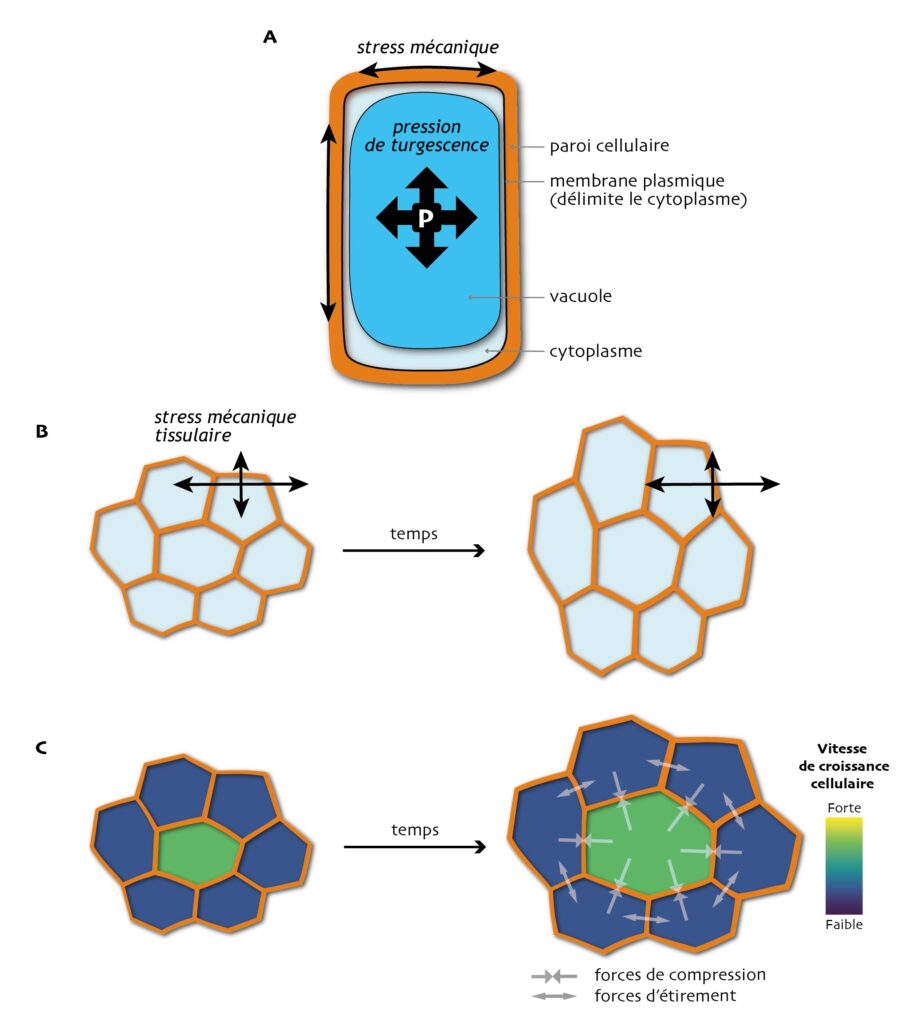
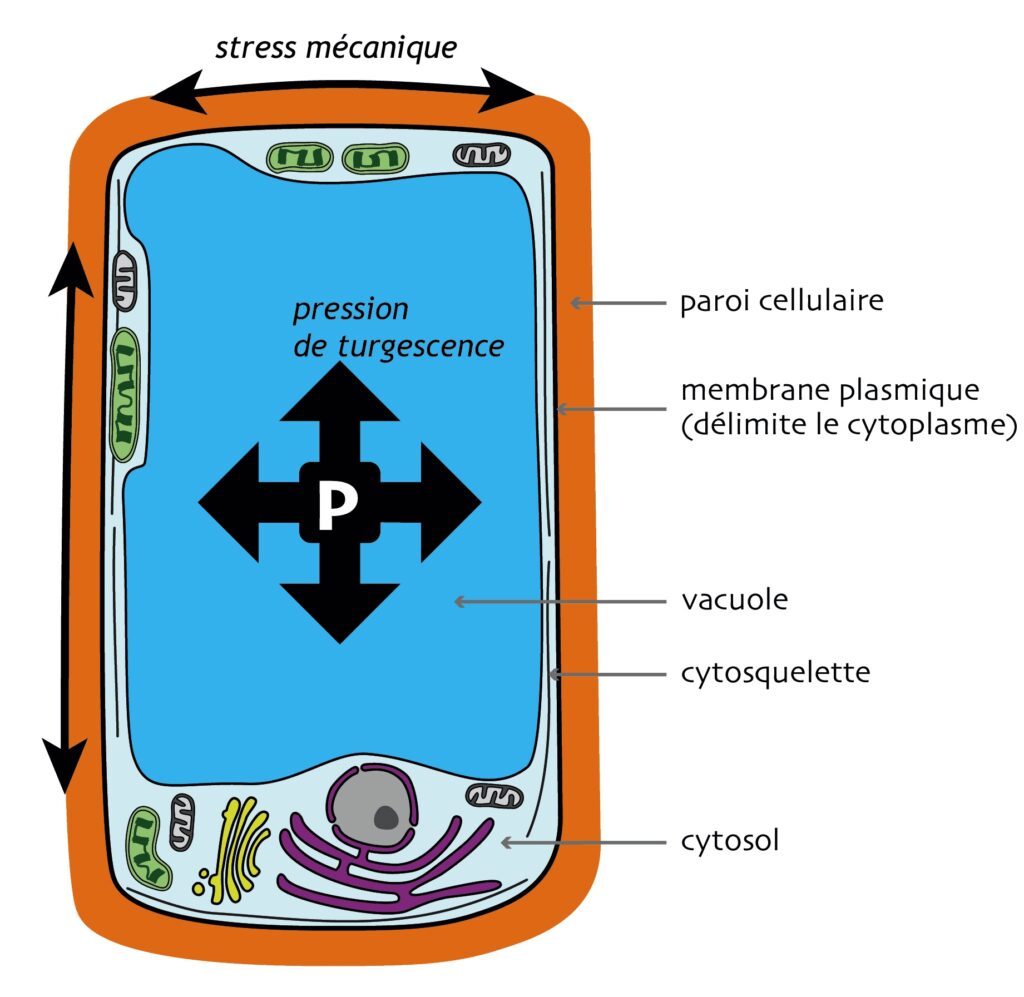
Dans ces vastes ensembles, les cellules végétales sont fermement maintenues solidaires les unes des autres par un matériau très épais et rigide, appelé paroi cellulaire, fait de différentes molécules (notamment des polymères de sucres). Cette contrainte particulière fait que le destin de chaque cellule est irrémédiablement lié à celui de ses voisines immédiates, desquelles elle ne pourra jamais se séparer de toute sa vie. Et, comme nous, les cellules végétales sont stressées ! Elles ne subissent pas un stress psychologique mais un « vrai » stress, c’est-à-dire un stress bio-mécanique. Et pas des moindres, puisqu’il est issu de l’équilibre fin entre la pression interne aux cellules qui peut atteindre plusieurs bars (soit la pression d’un pneu de camion) et les tensions qui s’accumulent dans les parois. En effet, la cellule végétale a la particularité de contenir un compartiment appelé vacuole qui se gonfle et se dégonfle et exerce une forte pression sur la paroi cellulaire (c’est la pression de turgescence). La paroi, elle, évite à la cellule d’éclater, comme une chambre à air dans un pneu, accumulant en elle une énorme tension mécanique (Figure 1A).
Paradoxalement, ces forces mécaniques sont le moteur de la croissance des cellules : sous l’effet du stress mécanique très fort, les parois, rigides mais tout de même un peu déformables, s’étirent petit à petit, permettant aux cellules de gagner en volume [1, 2].
Dans quelle direction pousser ? Le stress montre la voie
Les cellules « mesurent » le stress qu’elles subissent au niveau de leur paroi, et ajustent leur croissance en fonction. Le stress varie selon la direction (on dit qu’il est anisotrope) et, chez les plantes, les cellules tendent à croître dans la direction perpendiculaire à la direction du stress le plus fort. Ce principe directeur — un brin paradoxal — a un rôle central dans la mise en forme (la morphogenèse) des plantes, car les « champs de stress mécaniques » qui parcourent les tissus influencent grandement la direction dans laquelle les cellules s’étirent et se divisent en priorité, déterminant la forme que va acquérir l’organe (Figure 1B) [3, 4].
Mais chaque cellule a malgré tout une part de liberté : elle ne croît pas tout à fait au même rythme, ni dans la même direction, ni ne se divise au même moment que ses voisines immédiates. Comme elles sont « collées » les unes aux autres, les cellules adjacentes exercent les unes sur les autres des forces additionnelles, qui influent sur leur développement individuel (Figure 1C). Difficile alors de prédire comment un tel système, dit complexe, va évoluer au cours du temps. En 2012, une équipe internationale de biologistes, rassemblée autour de Magalie Uyttewaal, alors à l’École Normale Supérieure de Lyon, s’est demandé comment le comportement individuel des cellules évoluait dans un tissu en croissance sous l’effet du stress : les différences naturelles entre cellules sont-elles accentuées, ou au contraire gommées ?
Face au stress, des cellules (virtuelles) grandissent à différentes allures
Pour répondre à ces questions, les chercheur·e·s ont construit un modèle numérique de tissu végétal. Pourquoi un modèle numérique ? Parce qu’il est très difficile de suivre, sur une plante réelle, la vitesse de croissance de cellules individuelles au cours du temps. De plus, il est impossible d’isoler chaque cellule pour voir quelle vitesse de croissance propre elle adopterait sans contrainte, hors du tissu (ce que les auteur·e·s de l’étude appellent croissance autonome). De même, le stress mécanique subi par chaque cellule et sa direction privilégiée ne peuvent être mesurées directement ; on ne peut qu’estimer une valeur et une anisotropie moyenne, à l’échelle locale, selon la géométrie du tissu et la pression de turgescence moyenne des cellules (elle-même mesurée indirectement !). Enfin, un modèle numérique permet de mieux comprendre le fonctionnement d’un système complexe (ici un organisme vivant) en le simplifiant, en réduisant le nombre de facteurs impliqués [*].
Le modèle de tissu construit par les chercheur·e·s est plat, à deux dimensions. C’est un ensemble de polygones (chacun représentant une cellule) dont les parois sont des segments, reliés les uns aux autres par des sommets (leurs « coins » ; Figure 2). Ce modèle et sa croissance reproduit les principales caractéristiques des tissus végétaux en croissance :
- les cellules, sous forte pression interne, croissent lentement au cours du temps par l’étirement de leur paroi ;
- elles restent collées entre elles par leur parois communes, sans possibilité de déplacement les unes par rapport aux autres ;
- chaque cellule a une vitesse et une direction « de consigne » propre, qu’elle aurait si elle était seule (la croissance autonome) ; celle-ci varie d’une cellule à l’autre de manière aléatoire ;
- chaque cellule subit donc un stress anisotrope, « ressenti » dans ses parois, qui résulte à la fois de sa propre pression de turgescence et des contraintes imposées par ses voisines immédiates qui la « tirent » ou la « poussent » dans différentes directions ;
- enfin, chaque cellule est sensible au stress mécanique perçu, selon la « règle » établie de longue date par les botanistes : elle tend à orienter sa croissance dans la direction où le stress est le plus faible, et ce d’autant plus que le stress et/ou son anisotropie sont élevés. Il s’agit d’une rétroaction mécanique (= le stress, généré par les cellules, agit en retour sur leur propre comportement), un paramètre clé du modèle que les auteur·e·s peuvent faire varier à volonté. On peut faire l’analogie avec nos propres existences où, face au stress, nous nous comportons et prenons des décisions de manière — plus ou moins consciente — à minimiser ce stress !
Les fortes contraintes mécaniques que les cellules exercent les unes sur les autres en grandissant, et leur ajustement individuel en réponse au stress, font que leur croissance effective diffère de leur croissance autonome. La Figure 2A montre ce que génère une simulation, selon qu’on introduit, ou pas, une rétroaction : pour deux tissus rigoureusement identiques au départ, le stress, son anisotropie et sa direction principale et, donc, les rythmes de croissances des cellules, sont tous différents. Les scientifiques réalisent un grand nombre de simulations informatiques de la croissance du tissu virtuel (comportant 100 cellules) avec différents degrés de rétroaction mécanique, et analysent ensuite statistiquement les résultats.
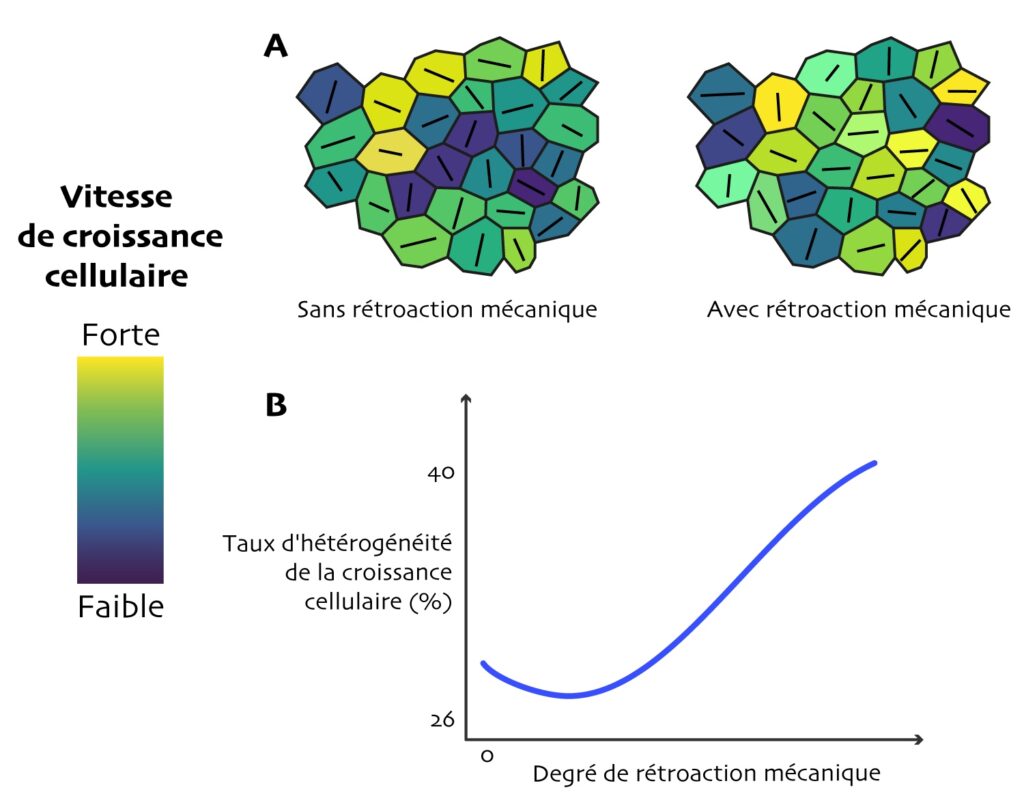
Surprise ! La rétroaction mécanique accroît les différences de vitesse de croissance entre cellules (Figure A2 : il y a davantage d’extrêmes de couleur à droite qu’à gauche ; Figure 2B) ! Autrement dit, l’hétérogénéité de la croissance, calculée pour chaque cellule comme la variabilité entre sa propre croissance et celle de toutes ses voisines immédiates, est promue par le stress. En fait, un tout petit peu de rétroaction rend la croissance entre cellules voisines plus homogène par rapport à pas de rétroaction du tout, mais cet effet n’est vrai que jusqu’à une certaine valeur-seuil (Figure 2B : le « creux » de la courbe bleue). Au-delà, l’hétérogénéité augmente de nouveau, et de manière presque linéaire avec le degré de rétroaction (Figure 2B : le milieu de la courbe est presque une droite). Ainsi, à l’image de la société humaine où, sous l’effet du stress subi par chaque personne, les parcours de vie de chacun prennent des directions divergentes et se différencient, les « trajectoires » de croissance de chaque cellule végétale se différencieraient sous l’effet du stress… mécanique. Cependant, les auteur·e·s ne se sont pas contentés de ce modèle très théorique. Elles et ils ont tenté d’observer, sur de vraies plantes, si cet effet du stress sur le comportement des cellules se confirme.
Et dans une vraie plante ? Plus il y a de stress, plus les écarts entre cellules se creusent !
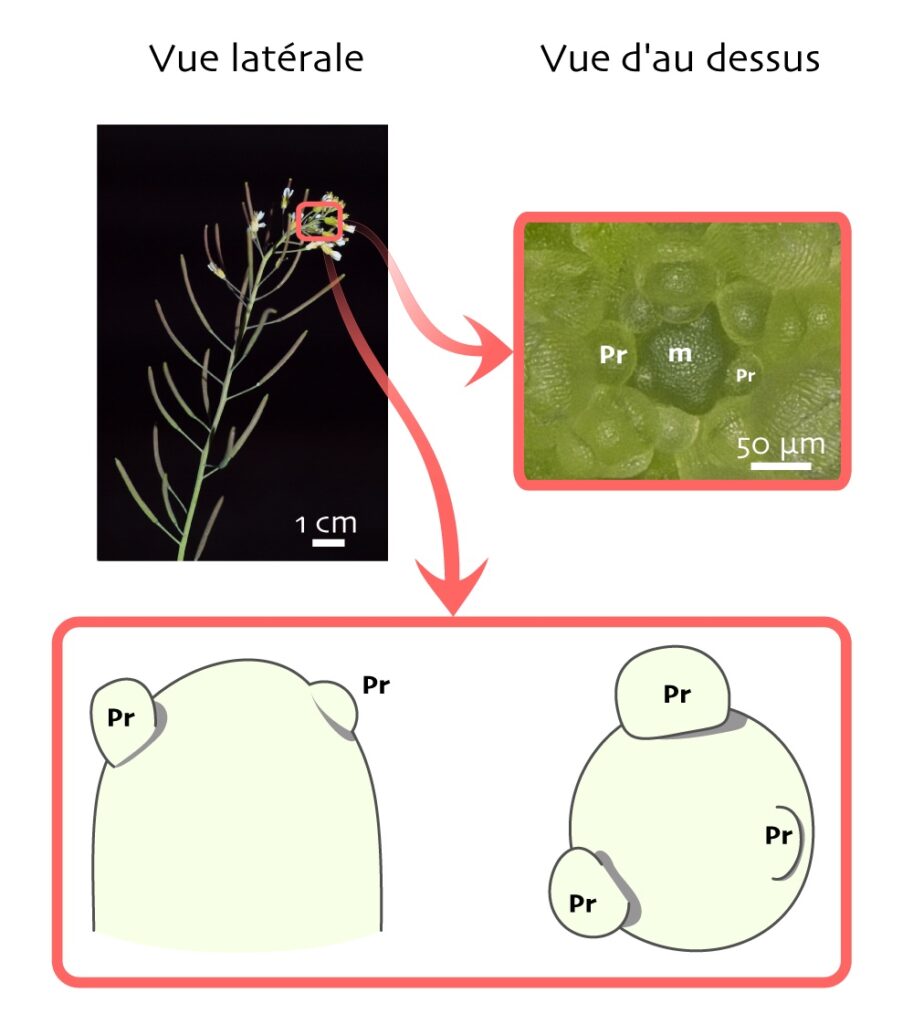
Pour cela, les scientifiques se sont tourné·e·s vers la plante star des laboratoires de biologie végétale : Arabidopsis thaliana, une petite plante de la famille du chou (Brassicaceae) [**] (Figure 3). En effet, on connaît chez cette plante, intensément étudiée depuis des décennies, des centaines de variétés présentant des altérations dans certains gènes (naturelles ou générées artificiellement en laboratoire) : ceux-ci peuvent être « sur-exprimés » ou « sous-exprimés », voire rendus non-fonctionnels. On appelle ces variétés des lignées mutantes. Ces mutants donnent l’opportunité d’étudier comment certains gènes interviennent dans divers processus physiologiques, comme la morphogenèse. Or, on connaît un mutant (que l’on nommera simplement « le mutant » ici) chez qui la fameuse rétroaction mécanique, par laquelle les cellules orientent leur croissance dans la direction du moindre stress, est diminuée ! Un peu comme si, dans notre société, les individus réagissaient moins au stress, se laissant moins contraindre par les aléas de l’existence. Si le modèle de tissu végétal décrit au-dessus est juste, on doit s’attendre à ce que la croissance cellulaire y soit plus homogène.
Nos biologistes ont cherché, plus précisément, à répondre à deux questions :
- Est-ce que l’hétérogénéité locale de croissance des cellules augmente avec l’intensité du stress et/ou son anisotropie ?
- Est-ce que ce lien est réduit quand la rétroaction mécanique est moins efficace ?
Pour cela, les chercheurs et chercheuses ont mesuré l’hétérogénéité de croissance cellulaire chez la plante sauvage (le « sauvage ») et le mutant. Comme cela a été dit plus haut, ce n’est pas une tâche simple. Les biologistes ont opté pour une méthode originale : elles et ils ont réalisé des moules en polymère de méristèmes [***] qui sont les extrémités des tiges où la morphogenèse est très active : c’est là que prennent naissance les organes aériens (ici, des fleurs) sous forme de petites protrusions appelées primordia (Figure 3). Deux moules de chaque méristème sont pris à un jour d’intervalle, puis observés au microscope électronique à balayage (MEB). On peut ainsi mesurer la vitesse de croissance des cellules (de combien leur aire s’est accrue en 24 h) et donc l’hétérogénéité du rythme de croissance entre cellules voisines, comme avec le modèle.
Globalement, la croissance entre cellules voisines est apparue plus hétérogène dans les méristèmes du sauvage que dans celui du mutant, ce qui confirme d’emblée la « prédiction » du modèle numérique : chez les plantes réelles, la rétroaction du stress mécanique sur la croissance favorise les écarts de croissance entre cellules, amplifiant donc le « chaos cellulaire » ! Cette rétroaction étant moins efficace chez le mutant, les croissances tendent à croître plus à l’unisson.
Par ailleurs, les mesures localisées de croissance cellulaire sur la surface des méristèmes a aussi permis de constater que l’hétérogénéité de croissance augmentait depuis le centre du méristème jusqu’à sa périphérie, atteignant un maximum aux « aisselles » des primordia floraux. C’est intéressant parce que, en raison de la topologie de surface du méristème, l’intensité du stress et son anisotropie augmentent justement depuis le centre vers sa périphérie et dans les aisselles des jeunes organes qui émergent à ce niveau-là [****]. L’hétérogénéité cellulaire augmente donc bien avec « l’intensité du signal », comme prédit par le modèle numérique (voir la courbe de la Figure 2B).
Un chaos cellulaire… utile à la morphogenèse ?
Cette étude a mis en évidence un phénomène curieux : dans les méristèmes des tiges de la petite plante Arabidopsis thaliana, le stress mécanique et son anisotropie, essentiels pour guider la morphogenèse, semble aussi accroître les inégalités de croissance entre cellules. Cela veut-il dire que la morphogenèse dans le méristème est peu efficace, limitée, « brouillée » par une sorte de chaos cellulaire que rien ne peut atténuer ?
À ce stade de leurs travaux, les chercheur·e·s n’avaient pas la réponse à cette question, mais ils ont proposé une hypothèse : ces écarts de croissance à petite échelle seraient en fait utiles à la morphogenèse. En effet, le mutant étudié ci-dessus, dont les auteur·e·s ont montré que la croissance cellulaire est localement plus homogène, présente aussi une morphogenèse défaillante : la surface du méristème est plus irrégulière et les jeunes primordia floraux en train d’émerger s’en « séparent » moins bien : les plis entre eux et le dôme central sont moins profonds et plus larges. Les auteur·e·s proposent donc que l’accentuation des écarts entre cellules proches permet de régulariser la géométrie des zones en croissance et facilite l’émergence des organes. Celle-ci requiert en effet que des cellules se mettent localement à croître (puis à se diviser) très rapidement, quand leurs voisines très proches, voire immédiates (avec lesquelles elles ont une paroi en commun) gardent une vitesse de croissance faible. Soit, une forte hétérogénéité locale. Des études ultérieures, menées par la même équipe semblent accréditer cette théorie… dans certaines conditions [5, 6].
Cette étude illustre aussi un thème récurrent en biologie : le comportement stochastique (= aléatoire) d’un grand nombre d’agents en interaction, provoque, à une échelle supérieure d’organisation, des phénomènes impossibles à prédire à partir de la simple somme de ces agents, et parfois même contre-intuitifs [7]. Depuis maintenant une vingtaine d’années, les scientifiques ont ainsi constaté que la stochasticité mécanique, en principe opposée à l’ordre et au contrôle qu’on imaginerait nécessaire dans le développement des êtres vivants, jouaient en fait un rôle très important dans la morphogenèse [8, 9]. Une complexité qu’on ne soupçonnerait pas, au premier abord, au cœur des plantes les plus humbles de nos jardins !
[*] Les modèles offrent aussi l’avantage de faire varier quantitativement les paramètres et de simuler informatiquement l’évolution du « système » avec ces valeurs. On peut alors mener des études statistiques précises, alors que de telles études sont beaucoup plus lourdes à mener sur des organismes vivants, voire impossibles si on ne peut modifier directement les paramètres.
[**] Appelée arabette des dames en français, c’est une petite plante herbacée de la même famille que les choux, le colza ou le radis. Elle est utilisée depuis des décennies comme modèle pour la recherche en biologie végétale car elle est très facile à cultiver et se développe rapidement : on passe de la graine à la plante adulte et fleurie en seulement deux mois. Elle a aussi un petit génome, entièrement séquencé en 2000, dans lequel il est facile d’induire des mutations et d’insérer des gènes étrangers (transformation génétique), ce qui la rend idéale pour étudier les gènes et leurs fonctions.
[***] Les méristèmes, à l’extrême pointe des tiges et des racines, sont des zones à croissance active, avec en leur centre un amas de cellules indifférenciées (dites pluripotentes), responsable de l’élongation des tiges et des racines. Les méristèmes de tiges (méristèmes caulinaires, souvent lovés au cœur d’une rosette de jeunes feuilles), produisent aussi les différents organes aériens de la plante (feuilles, fleurs, etc.), qui apparaissent dans un ordre précis.
[****] Pour aller plus loin : La tige et le méristème peuvent être assimilés à un ballon sous pression, de forme cylindrique et terminé par un dôme. Or, les lois de la mécanique veulent que le stress dans les parois du ballon soit réduit et isotrope au sommet du dôme, puis qu’il augmente et devienne anisotrope à mesure qu’on se rapproche de la partie cylindrique (soit la périphérie du méristème, puis la tige). Dans cette dernière, le stress est deux fois plus élevé en circonférence (direction circonférentielle) que dans la direction parallèle à l’axe du cylindre (direction méridionale). À l’aisselle des primordia, la surface a topologiquement une forme de « selle à cheval » (la surface est convexe en direction circonférentielle et concave en direction méridionale), ce qui fait que l’anisotropie de stress devient localement très élevée : en direction méridionale il est devenu un stress de compression, qui tend à « écraser » la paroi alors que dans la direction circonférentielle, le stress (dit de tension ou d’étirement) tend à la déchirer !
[1] Geitmann A., et al., Mechanics and modeling of plant cell growth. Trends in Plant Science, 2009. DOI : 10.1016/j.tplants.2009.07.006. [Review]
[2] Harold M.F., To shape a cell: an inquiry into the causes of morphogenesis of microorganisms. Microbiological Reviews, 1990. DOI : 10.1128/mr.54.4.381-431.1990. [Review]
[3] Hamant O., et al., Developmental patterning by mechanical signals in Arabidopsis. Science, 2008. DOI : 10.1126/science.1165594. [Publication scientifique]
[4] Landrein B., et al., How mechanical stress controls microtubule behavior and morphogenesis in plants: History, experiments and revisited theories. Plant Journal, 2013. DOI : 10.1111/tpj.12188. [Review]
[5] Burian A., et al., A correlative microscopy approach relates microtubule behaviour, local organ geometry, and cell growth at the Arabidopsis shoot apical meristem. Journal of Experimental Botany, 2013. DOI : 10.1093/jxb/ert352. [Publication scientifique]
[6] Hong L., et al., Variable cell growth yields reproducible organ development through spatiotemporal averaging. Developmental Cell, 2016. DOI : 10.1016/j.devcell.2016.06.016. [Publication scientifique]
[7] Oates A.C., What’s all the noise about developmental stochasticity? Development, 2011. DOI : 10.1242/dev.059923. [Review]
[8] Hamant O., Shoot meristem progenitors emerge from mechanical heterogeneities. Developmental Cell, 2022. DOI : 10.1016/j.devcel.2022.08.004. [Courte note dans une revue académique]
[9] Hamant O., La troisième voie du vivant, Odile Jacob, 2022 [Livre grand public]
Publié le 20/10/2024
 Hervé Rabillé/Papier-Mâché/CC BY-NC-SA 4.0 2024
Hervé Rabillé/Papier-Mâché/CC BY-NC-SA 4.0 2024Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.
Ou comment se développe la forme des plantes
Écriture : Hervé Rabillé
Relecture scientifique : Cécile Feuillie et Éléa Hébérlé
Relecture de forme : Charlotte Rémy et Eléonore Pérès
Temps de lecture : environ 18 minutes.
Thématiques : Développement, Biologie végétale, Biologie cellulaire (Biologie)
Publication originale : Uyttewaal M., et al., Mechanical stress acts via katanin to amplify differences in growth rate between adjacent cells in Arabidopsis. Cell, 2012. DOI : 10.1016/j.cell.2012.02.048. Accès libre sur HAL (pdf).
Au sommet des tiges des plantes, de nouveaux organes aériens aux formes précises apparaissent en continu. Mais la croissance d’un grand nombre de cellules végétales s’accompagne d’irrémédiables conflits mécaniques, car elles sont entourées d’une solide paroi qui les lient les unes aux autres. En 2012, des chercheur·e·s ont montré, dans la petite plante Arabidopsis thaliana, que le stress mécanique accentue les écarts de croissance entre cellules voisines au lieu de les atténuer. Plus surprenant encore, cela semble faciliter le développement des formes de la plante !
Les plantes, comme tous les êtres vivants, ont des formes précises et reproductibles. C’est par la forme de leur port, de leurs tiges, de leurs feuilles et de leurs fleurs que l’on reconnaît, en premier lieu, les espèces végétales que l’on rencontre dans notre environnement — pour peu que l’on s’y intéresse. Bien sûr, ces formes n’apparaissent pas spontanément mais s’acquièrent progressivement au cours du développement : c’est ce qu’on appelle la morphogenèse [1].
Comment les plantes fabriquent-elles leurs organes ?
Les plantes terrestres (Embryophytes) sont multicellulaires, comme les animaux ou la plupart des champignons : elles sont composées de nombreuses cellules, regroupées en tissus, eux-mêmes rassemblés en organes avec une forme déterminée assurant une fonction (par exemple la feuille, qui produit les sucres par photosynthèse). Les propriétés et comportements individuels des cellules vont grandement influencer la manière dont se fait la morphogenèse à toutes les échelles d’organisation, notamment car les cellules modifient les propriétés mécaniques du tissu et celles-ci, en retour, affectent le comportement des cellules.
En termes physiques, une cellule est une goutte de matière visqueuse — le cytoplasme — délimitée par une membrane plasmique riche en lipides, éventuellement renforcée par des armatures moléculaires internes — le cytosquelette — et externes — la matrice extracellulaire (Figure 1). Chez les plantes, cette matrice est très épaisse et rigide, et appelée de ce fait paroi cellulaire. Elle impose une forme définie aux cellules et les maintient irrémédiablement attachées entre elles, sans possibilité de migration, contrairement à ce qui se passe dans les tissus animaux. Par ailleurs, la pression dans les cellules végétales est particulièrement forte : elles présentent un compartiment interne, la vacuole, qui « pompe » l’eau externe et génère ainsi une pression dite de « turgescence », parfois aussi élevée que celle d’un pneu de camion (soit plus de 5 bar) ! Cette turgescence soumet la paroi et, au-delà, tout le tissu à des stress mécaniques très forts (Figure 1), qui sont nécessaires pour « étirer » et, donc, faire croître toute la structure [2]. Difficulté supplémentaire : si chaque cellule tend à pousser à une vitesse un peu différente de ses voisines, alors elle génère sur ses consœurs des stress mécaniques additionnels [3]. Il s’agit là d’une contrainte très forte sur la morphogenèse des plantes, qui repose quasi uniquement sur la localisation et l’orientation, dans l’espace, de la croissance et de la division cellulaires [2]. Une tige, par exemple, se forme par la croissance de toutes ses cellules dans une direction principale, alors que les folioles d’une feuille composée naissent de petits groupes de cellules au bord du limbe qui se mettent à croître et à se diviser activement, quand la croissance de leurs sœurs d’à côté, elle, reste lente. Dans le premier cas, la croissance entre cellules voisines est relativement harmonisée : elles grandissent à une vitesse proche les unes des autres. La croissance est homogène, et il y a — en principe — peu de conflits mécaniques entre cellules voisines. Dans le second cas, en revanche, des cellules très proches grandissent à des vitesses très différentes : la croissance est localement hétérogène, et d’importants stress mécaniques peuvent surgir.
Un problème épineux pour les spécialistes du développement des plantes est donc de savoir comment les plantes parviennent à générer des organes aux formes précises et reproductibles avec un tel « jeu de mécano » cellulaire !
Le méristème apical, « haut-lieu » de la morphogenèse végétale
La partie idéale de la plante pour étudier les liens qui unissent stress mécanique, croissance des cellules et morphogenèse des organes, c’est le méristème apical caulinaire. C’est en effet là, à l’extrémité des tiges ou au cœur des rosettes de feuilles, que se forment tous les organes aériens (tiges, feuilles, bourgeons, fleurs, etc.). Le cœur du méristème est un tissu composé de petites cellules indifférenciées, et est lui-même divisé en deux zones : une zone centrale et une zone périphérique (qu’on abrégera ZC et ZP, Figure 2A). Dans la ZC, la croissance des cellules est lente et égale dans toutes les directions : elle est dite isotrope. Dans la ZP, la croissance s’accélère un peu et devient anisotrope : les cellules s’allongent davantage dans la direction méridionale que dans la direction azimutale [4] [*] — ici s’amorce déjà l’élongation de la tige. C’est aussi dans la ZP que les autres organes apparaissent, dans un ordre spatio-temporel précis, sous forme de primordia (singulier : primordium, Figure 2A), des « protrusions » dans lesquelles les cellules se mettent à grandir et à se diviser activement.
Le méristème a été bien étudié chez Arabidopsis thaliana, une petite plante herbacée [**]. Avec des techniques de microscopie, de biophysique et de génétique, les biologistes ont observé que le stress mécanique tissulaire y régule la direction et le rythme de croissance des cellules. En effet, des éléments du cytosquelette, les microtubules corticaux [***] (qu’on appellera simplement microtubules ici, pour simplifier) s’alignent dans la direction où le stress est le plus fort (Figure 2B-D) [4, 5 et 6]. Or, des microfibrilles de cellulose se déposent dans la paroi dans la même direction que les microtubules. Ces microfibrilles, très solides et rigides, agissent comme des câbles d’acier qui restreignent la croissance des cellules dans la direction où elles sont déposées (Figure 2C), contraignant les cellules à s’allonger principalement dans la direction perpendiculaire. Ainsi, dans le méristème, une chaîne d’instructions part d’un « signal primaire », le stress mécanique tissulaire, et aboutit à l’élongation anisotrope des cellules dans la direction où, paradoxalement, le stress mécanique est le plus faible. Cette voie de régulation est très importante dans le développement des plantes (Figure 2A-D) [4].
Mais alors, si le stress mécanique régule la direction de croissance des cellules, est-ce qu’il peut aussi réguler leur vitesse, et donc l’hétérogénéité de la croissance dans le tissu ? Et, si oui, quelle importance cela a-t-il dans l’émergence des organes ?
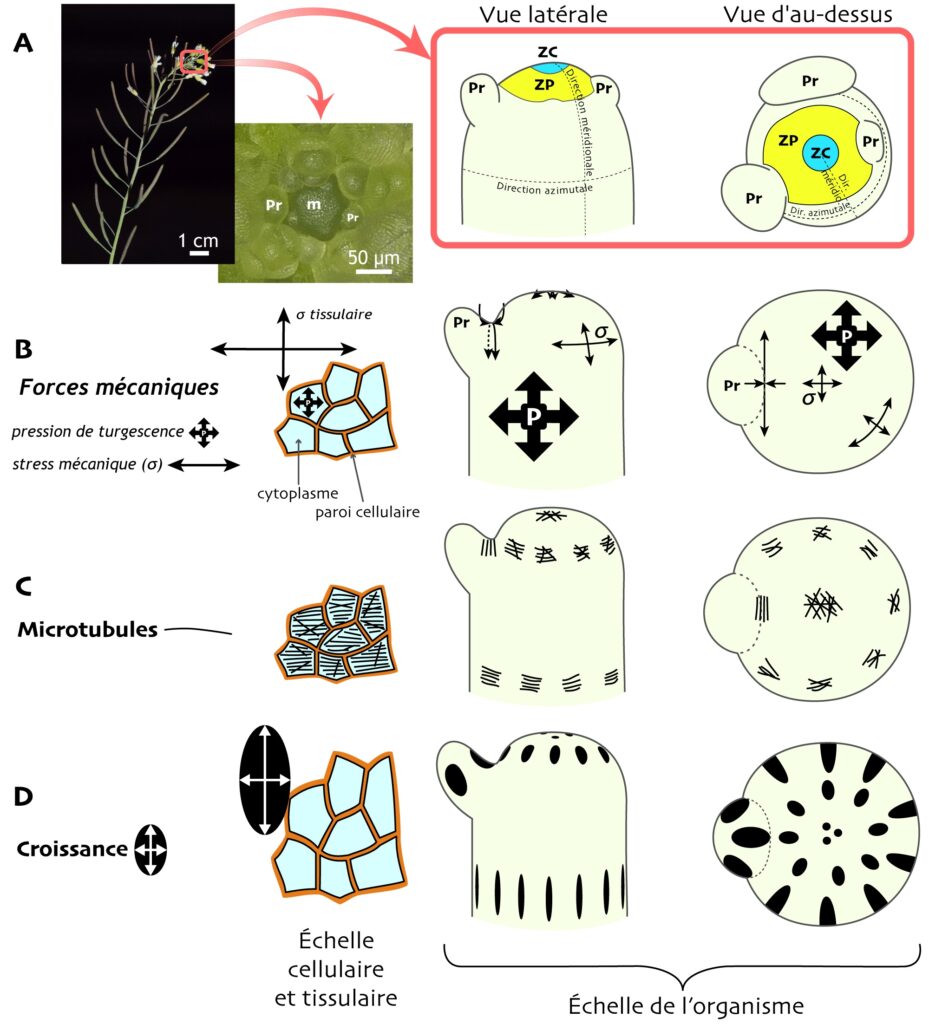
Un mutant d’Arabidopsis à la morphogenèse déficiente
Pour répondre à ces questions, Magalie Uyttewaal et ses collègues, auteur·e·s de l’étude qui nous intéresse ici, ont caractérisé une lignée mutante d’Arabidopsis qui a une morphogenèse anormale (on l’appellera simplement « le mutant » dans le reste du texte ; Figure 3). Cette lignée porte une mutation dans le gène codant une protéine appelée la katanine [7]. Normalement, celle-ci est capable de découper les microtubules. Mais la mutation a rendu la protéine non fonctionnelle : elle est incapable de couper les microtubules et ceux-ci se remodèlent donc moins vite que chez la plante sauvage (la plante non mutée, et qu’on appellera seulement « le sauvage »).
Par une série d’observations aux microscopes électronique à balayage (MEB) et à fluorescence, et des micromanipulations du méristème [****], l’équipe de recherche a confirmé le lien entre l’anisotropie du stress mécanique et la direction d’alignement des cellules. La mutation de la katanine rend les cellules effectivement moins capables de percevoir la direction principale du stress : leurs microtubules se réalignent moins dans la direction maximale du stress (Figure 3B) et gardent une distribution plus aléatoire (Figure 3C). Cela se traduit par une croissance du tissu plus isotrope (Figure 3D), ce qui explique en partie la forme anormale du méristème chez ce mutant (Figure 3A).Les biologistes se sont alors posé la question suivante : les défauts morphogénétiques du mutant pourraient-ils aussi résulter d’écarts de croissance plus importants, ou plus faibles, entre cellules proches au sein du tissu ?
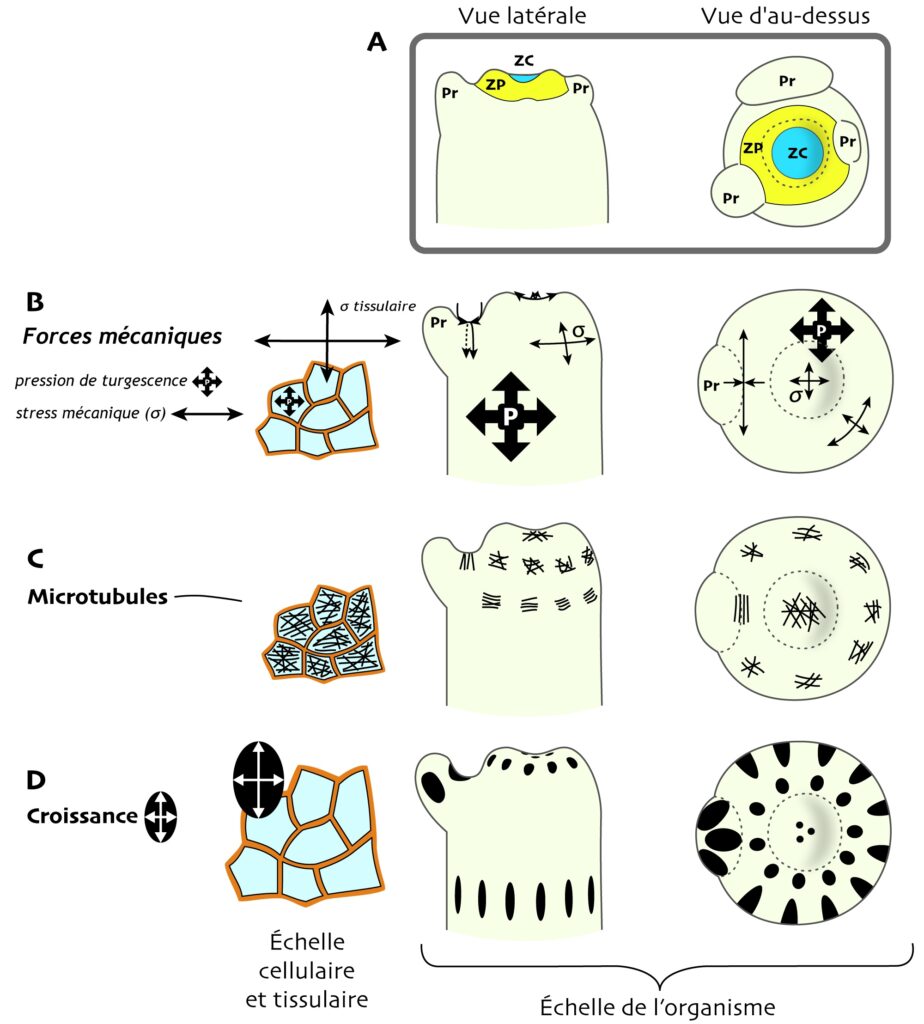
Comment le stress mécanique influe-t-il sur les écarts de croissance entre cellules ?
Hélas, sur une plante réelle il est très difficile de quantifier précisément l’effet du degré de sensibilité des cellules au stress mécanique sur leur dynamique de croissance. Le stress mécanique, par exemple, ne peut pas être mesuré expérimentalement dans le tissu, mais seulement calculé de manière indirecte et approchée. Pour contourner cette limitation, les chercheur·e·s ont donc construit un modèle numérique (dit in-silico). Avec l’essor de l’informatique, les modèles numériques sont devenus incontournables pour tester des hypothèses sur des systèmes physiques complexes et difficiles, voire impossibles à manipuler dans la réalité, y compris les organismes vivants.
Leur modèle reproduit un tissu végétal plat, en deux dimensions [*****], composé de « cellules » virtuelles qui sont en fait des polygones dont les parois sont des segments reliés les uns aux autres par des sommets (leurs « coins » ; Figure 4). La croissance des cellules est due à l’étirement de ces segments au cours du temps sous l’action de forces mécaniques.
Dans le modèle, chaque cellule a une vitesse et une direction de consigne, c’est-à-dire la vitesse et la direction qu’elle afficherait si elle était seule, sans contact avec d’autres cellules (Figure 4A), ce que les auteur·e·s appellent croissance autonome. Celle-ci a un caractère stochastique : elle diffère d’une cellule à l’autre, avec un certain degré de variabilité que l’on peut ajuster dans le modèle.
Mais leur vitesse de croissance effective diffère de leur croissance autonome car elles se gênent les unes les autres. Dans la Figure 4B, la cellule au milieu tend à grandir bien plus vite que ses voisines. En conséquence, elle les « pousse » (forces de compression) et génère chez elles, en réaction, des forces d’étirement. Ces cellules périphériques vont donc croître plus vite, et la cellule centrale, au contraire, va être ralentie. Tout cela se traduit aussi par un stress mécanique plus ou moins fort et anisotrope, propre à chaque cellule (Figure 4B, σ).Les cellules virtuelles sont sensibles à ce stress mécanique : elles orientent leur croissance prioritairement dans la direction de stress minimum. Ce degré de « sensibilité », appelé rétroaction mécanique, peut aussi être ajusté dans le modèle : plus il est élevé, plus les cellules ralentissent leur croissance dans la direction de stress maximum. La Figure 5A montre ce que génère une simulation selon qu’on introduit, ou pas, un certain degré de rétroaction : pour deux tissus rigoureusement identiques au départ, la vitesse de croissance, la valeur et la direction de stress maximal sont tous différents, ce qui mènera à des profils de croissance différents. La sensibilité mécanique des cellules agit donc bien en retour sur le profil de stress mécanique et leur croissance (d’où le terme rétroaction).
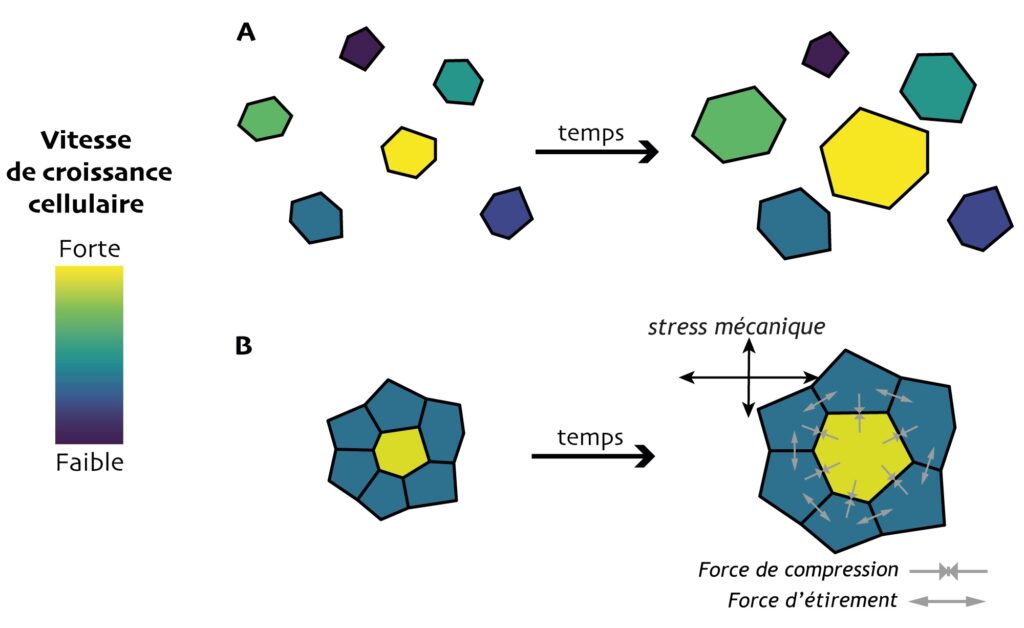
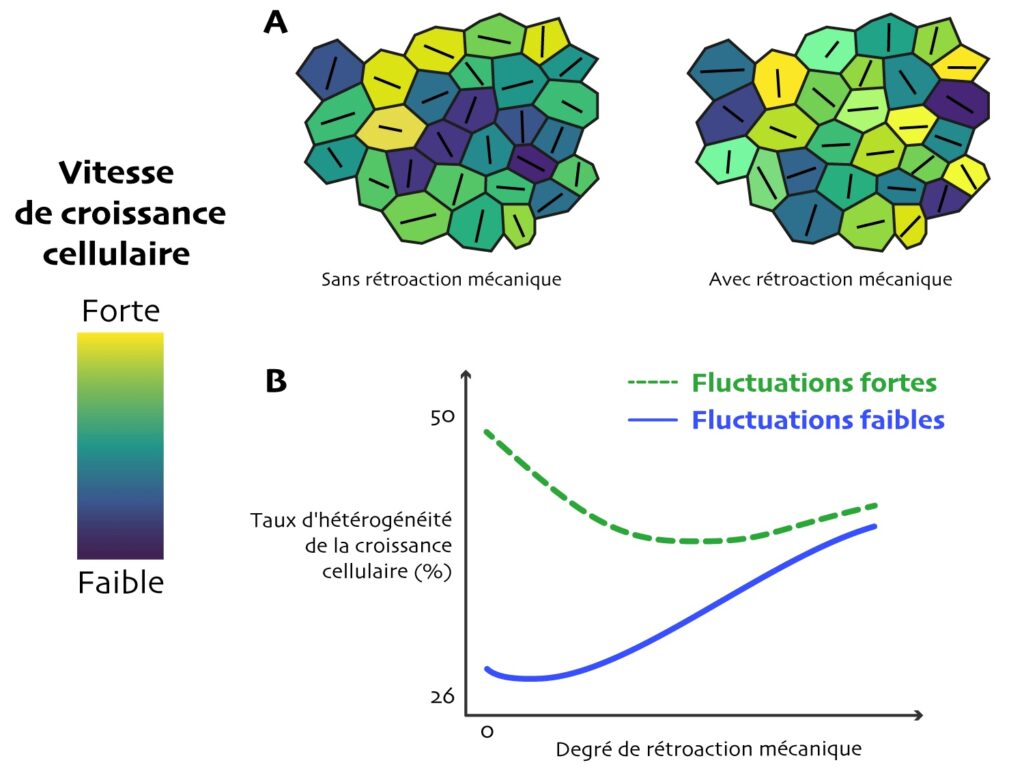
Pour voir si cette rétroaction harmonise la croissance entre voisines ou, au contraire, favorise la compétition, les chercheur·e·s ont fait des simulations informatiques de la croissance d’un tissu de 100 cellules avec différentes valeurs de stochasticité de croissance autonome et de rétroaction (160 simulations indépendantes pour chaque paire de valeur). L’hétérogénéité des paramètres cellulaires est calculée pour chaque cellule par rapport à ces voisines immédiates, puis moyennées sur l’ensemble du tissu.
L’analyse statistique des résultats de simulations révèle une surprise : la rétroaction mécanique accroît généralement les différences de vitesse de croissance entre cellules (Figure 5B) ! Un tout petit peu de rétroaction rend la croissance entre cellules voisines un peu moins hétérogène par rapport à pas de rétroaction du tout (parties gauches des courbes en Figure 5B), mais cet effet n’est vrai que jusqu’à une certaine valeur (hétérogénéité minimum, creux des courbes). Au-delà, l’hétérogénéité augmente de nouveau, et de manière presque linéaire avec la rétroaction (parties droites des courbes). Plus surprenant encore : quand les cellules tendent à croître à une vitesse proche les unes des autres (fluctuation stochastique faible, Figure 5B, courbe bleue), la rétroaction rend la croissance entre cellules presque toujours plus hétérogène !
Au vu de ce résultat, les auteur·e·s ont suggéré que, dans les plantes réelles, le contrôle exercé par l’anisotropie du stress mécanique sur la croissance des cellules pourrait favoriser les écarts de croissance entre cellules voisines. Le mutant de la katanine permet justement de le vérifier expérimentalement. En effet, chez celui-ci les cellules perçoivent moins le stress mécanique par rapport au sauvage : la rétroaction mécanique est plus faible. On peut donc s’attendre à ce que la croissance cellulaire y soit plus homogène.
La croissance cellulaire est localement plus homogène chez le mutant de la katanine
Pour le vérifier, les chercheur·e·s ont mesuré l’hétérogénéité de croissance cellulaire chez le sauvage et le mutant à partir de moules en polymère de méristèmes, réalisés à 24 h d’intervalle et observés en MEB. Sur toute la surface du méristème (du centre jusqu’aux aisselles des primordia de fleurs), les biologistes ont mesuré de combien s’est étendue l’aire de chaque cellule entre les deux dates d’observation, et de là ont calculé sa vitesse d’expansion. Les biologistes ont alors calculé l’hétérogénéité du rythme de croissance entre cellules voisines, comme dans le modèle décrit ci-dessus, et ont observé comment celle-ci évolue dans l’espace. Connaissant la distribution et l’orientation théorique du stress sur la même zone (Figures 2 et 3), les biologistes ont regardé si hétérogénéité et stress mécanique évoluent conjointement.
Globalement, la croissance entre cellules voisines est apparue plus hétérogène dans le méristème de la plante sauvage que dans celui du mutant. De plus — et même si les données étaient très variables d’un méristème à l’autre — l’hétérogénéité de croissance augmente depuis le centre du méristème jusqu’à sa périphérie… comme le fait le stress mécanique, qui augmente en intensité et en anisotropie du cœur du méristème vers le bord (Figure 2B). Le degré d’hétérogénéité semble donc augmenter avec l’intensité du « signal mécanique ». Chez le mutant, la même tendance est observée, mais elle reste plus faible (Figure 6).
La différence entre le mutant et le sauvage se creuse… au niveau des plis qui séparent les primordia, en bordure du méristème. La topologie de ces régions très étroites rend le stress mécanique très anisotrope (Figure 2B). Dans la plante sauvage, l’hétérogénéité de croissance y est très importante, avec des différences de croissance entre cellules adjacentes entre 400 et 800 % environ. Elle augmente aussi chez la plante mutante, mais reste faible comparé au sauvage, car elle n’y est plus que d’environ 29 à 43 % (Figure 6).
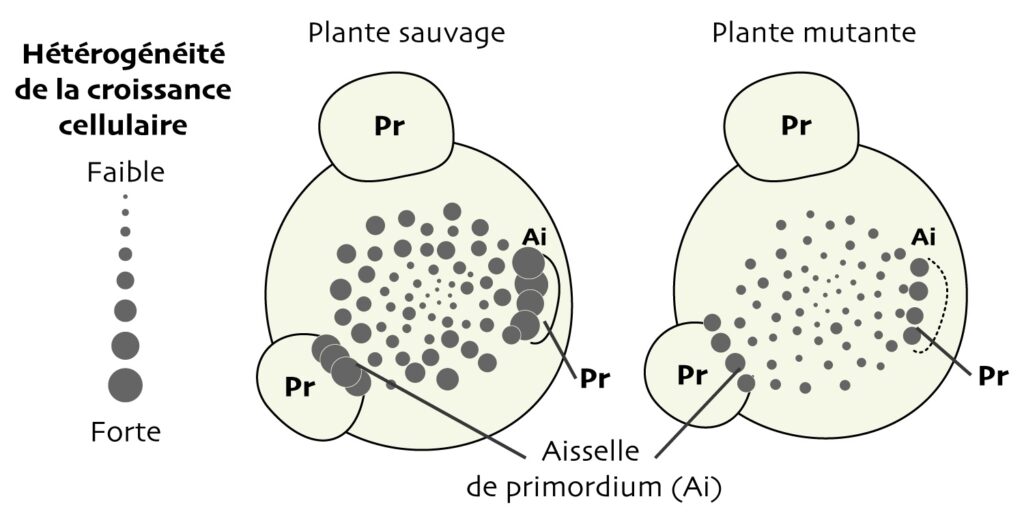
Ces résultats suggèrent que, chez le mutant, la perte de sensibilité des cellules à l’anisotropie de stress mécanique conduit à une homogénéisation de la croissance entre cellules voisines. Et donc que, chez le sauvage, le stress promeut bien cette hétérogénéité !
Mais en quoi cela est-il « utile » à la plante ?
Creuser les écarts de croissance entre cellules pour une morphogenèse plus précise
Dans le méristème apical d’Arabidopsis, le stress mécanique semble accentuer l’hétérogénéité de croissance entre cellules voisines, via la réorganisation des microtubules. Cela veut-il dire que la morphogenèse dans le méristème est peu efficace, limitée par une sorte de « chaos cellulaire » que rien ne peut contrecarrer ?
À l’opposé de ce scénario « pessimiste », les auteur·e·s proposent que ce phénomène serait, au contraire, utile à la plante : il favoriserait l’émergence de l’organe et sa démarcation vis-à-vis du reste du méristème, où la croissance reste beaucoup plus modérée.Uyttewaal et ses collègues proposent le scénario suivant : lorsqu’un primordium émerge en périphérie du méristème, la croissance commence à accélérer dans cette zone et induit un stress mécanique additionnel tout autour du primordium. Cela exacerbe l’anisotropie du stress mécanique dans la direction azimutale (Figure 7A et D). Dans la file de cellules où l’anisotropie est maximale, les microtubules s’alignent massivement dans la direction azimutale et la croissance dans cette direction est bloquée (Figure 7B et E). Les cellules immédiatement adjacentes, notamment celles à la base du futur primordium, sont, elles, moins entravées et peuvent grandir plus vite (Figure 7B, cellules à droite). Leur croissance accentue encore le stress mécanique dans la file centrale de cellules et prolonge le blocage de leur croissance (Figure 7C). Rapidement, le tissu commence localement à se « plier » car les cellules bloquées ne croissent pas, ou peu ; ce pli marque une frontière physique plus nette entre le primordium et le méristème, deux régions aux destins développementaux très différents (Figure 7C et F). En 2013, la même équipe a publié une autre étude expérimentale qui semble appuyer ce scénario [8].
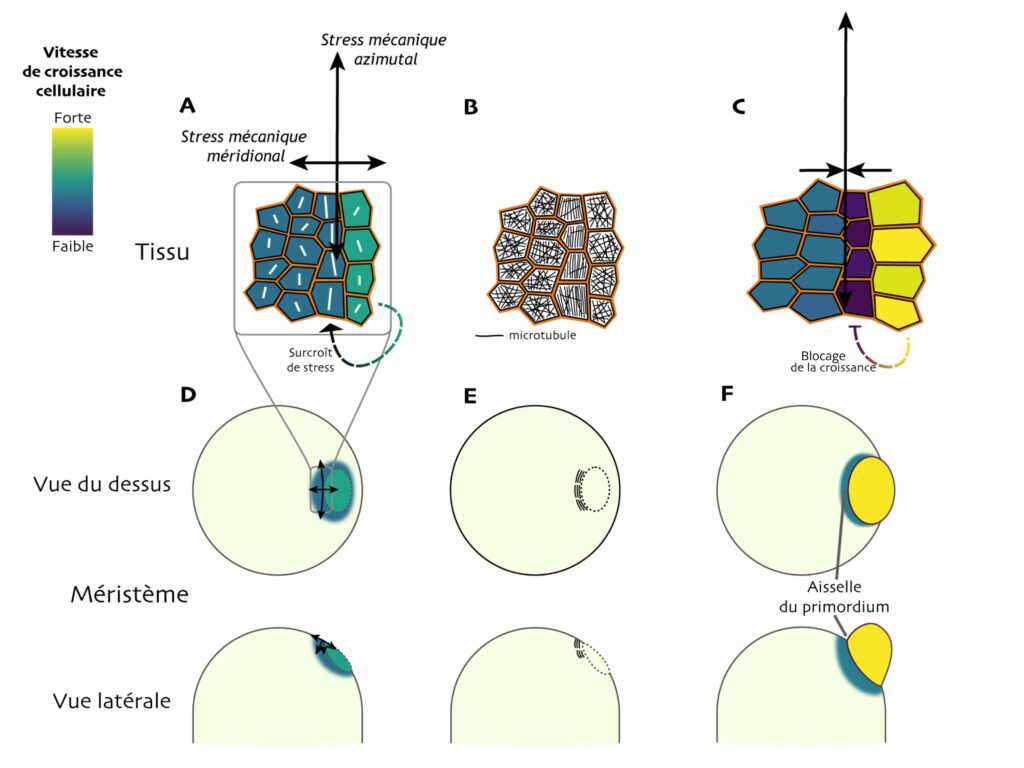
Ainsi, en favorisant la variation de croissance entre cellules juxtaposées, la rétroaction du stress mécanique sur les cellules favoriserait la précision de la morphogenèse tissulaire à de petites échelles spatiales — de l’ordre de quelques cellules. Chez le mutant, l’absence de katanine rendrait ce phénomène moins efficace : la croissance cellulaire plus homogène retarderait la pliure du tissu à la base des primordia. Reste à savoir, cependant, comment cela affecte le développement des organes à des stades plus tardifs.
Un mécanisme… pas du tout universel
Cette théorie du stress comme « amplificateur du chaos cellulaire » n’est pas vérifiée partout, loin de là. Des contre-exemples sont connus, y compris au sein d’Arabidopsis ! Par exemple, l’accumulation locale de stress dans un tissu en expansion peut rapidement bloquer la croissance cellulaire dans tout le tissu. C’est ce qui est observé dans l’aile en formation de la mouche du vinaigre, Drosophila melanogaster [9, 10]. Des travaux plus récents, réalisés sur Arabidopsis par la même équipe de l’ENS de Lyon, ont montré un effet similaire dans le sépale — un des organes de la fleur. Le stress s’accumule à la pointe du sépale pendant son développement, et y bloque la croissance des cellules. Contrairement au modèle proposé plus haut, cela ne favorise pas l’expansion des cellules situées plus bas, au contraire ! À un moment donné, le surcroît de stress se propage de proche en proche depuis la pointe jusqu’à la base du sépale, comme une vague « figeant » rapidement la croissance de toutes les cellules de l’organe, qui acquiert alors sa forme définitive [11].
Toujours dans le sépale, mais à une échelle plus petite, les cellules qui formeront plus tard des trichomes [******] croissent plus vite que leurs voisines. Mais la forte tension mécanique que celles-ci accumulent bloque en retour l’expansion des précurseurs de trichomes, restreignant au final les différences de taille cellulaire [12]. En fait, Uyttewaal et collègues ont pressenti, dès 2012, que la manière dont le stress mécanique régulait le comportement des cellules dépendait du contexte développemental où il s’exerce — nature de l’organe, stade de développement, topologie, taille, etc.
Quoi qu’il en soit, toutes ces études — et bien d’autres encore — confirment que le stress mécanique tissulaire, souvent généré par le caractère stochastique du comportement des cellules, peut servir de « signal régulateur » avec un rôle avantageux dans le développement [13, 14] ! Qui aurait pu croire que la pousse des petites herbes de nos jardins, en apparence si simples et anodines, recelait une telle complexité ?
[*] Prenez n’importe quel point à la surface du méristème et de la tige : la direction méridionale est celle qui passe par ce point et le centre du méristème. La direction azimutale, elle, est perpendiculaire à la première, et trace un cercle autour du centre du méristème.
[**] Arabidopsis thaliana, appelée arabette des dames en français, est une petite plante herbacée de la famille des Brassicacées (dans laquelle on trouve également les choux, le colza, le radis…). Elle est utilisée depuis des décennies comme modèle pour la recherche en biologie végétale, car elle est très facile à cultiver, se développe rapidement (elle peut compléter tout son cycle de vie en seulement deux mois). Elle a aussi un petit génome, entièrement séquencé en 2000, dans lequel il est facile d’induire des mutations et d’insérer des gènes étrangers (transformation génétique), ce qui la rend idéale pour étudier les gènes qui contrôlent le métabolisme, la physiologie et le développement.
[***] Chez les plantes, les microtubules corticaux sont un ensemble de microtubules situés sous la membrane plasmique des cellules, parallèle à celle-ci, formant une sorte de maille. Au sein de cette maille, les microtubules peuvent avoir une orientation plus ou moins aléatoire, ou au contraire être alignés parallèlement les uns aux autres, voire rassemblés en faisceaux.
[****] La micromanipulation consiste à exercer des forces sur l’organe à l’échelle du micromètre ou de la dizaine de micromètre, avec des outils adaptés, et à observer, au microscope, la réponse des tissus et des cellules. En mesurant le degré de déformation en fonction de la force appliquée ou induite, les biophysicien·ne·s sont capables de calculer la valeur approximative de certaines propriétés mécaniques de l’objet (élasticité, pression interne). Dans la présente étude, les chercheur·e·s ont effectué trois grands types de manipulation : 1) compression des méristèmes (préalablement débarrassés des jeunes feuilles qui les entouraient) entre deux microplaques, pour estimer l’élasticité globale du tissu et sa pression interne ; 2) destruction de petits groupes de 3-4 cellules au laser pour reconfigurer le stress mécanique de surface dans les cellules adjacentes ; 3) cartographie des propriétés mécaniques du tissu en surface au microscope à force atomique — une pointe montée sur un cantilever avec, au bout, une bille de 5 µm de diamètre qui appuie sur le tissu, et dont l’enfoncement en fonction de la force exercée est mesuré très précisément avec un laser.
[*****] La courbure du tissu est ignorée dans ce modèle car, dans le méristème, elle est relativement faible à l’échelle d’une cellule unique, et son effet sur le profil local de stress est donc considéré comme négligeable. Cela autorise les scientifiques à ne modéliser la croissance cellulaire que dans 2 dimensions (la « surface » du tissu), ce qui simplifie grandement les calculs. Cependant, cette condition de faible courbure n’est plus tellement vraie au niveau des plis à la base des primordia en croissance. Comme tous les modèles, celui proposé par Uyttewaal et ses collègues repose donc sur un certain degré de simplification qui limite la portée de ses « prédictions ». Une extension intéressante du modèle serait de tester l’effet de la courbure sur la réponse des cellules.
[******] Petits poils qui poussent à la surface des organes aériens des plantes (feuilles, fleurs, fruits, etc.). Quand présents en abondance, ils peuvent donner à la surface une texture duveteuse.
[1] Harold M.F., To shape a cell: an inquiry into the causes of morphogenesis of microorganisms. Microbiological Reviews, 1990. DOI : 10.1128/mr.54.4.381-431.1990. [Review]
[2] Geitmann A., et al., Mechanics and modeling of plant cell growth. Trends in Plant Science, 2009. DOI : 10.1016/j.tplants.2009.07.006. [Review]
[3] Uyttewaal M., et al., Integrating physical stress, growth, and development. Current Opinion in Plant Biology, 2010. DOI : 10.1016/j.pbi.2009.10.004. [Review]
[4] Hamant O., et al., Developmental patterning by mechanical signals in Arabidopsis. Science, 2008. DOI : 10.1126/science.1165594. [Publication scientifique]
[5] Hamant O., Widespread mechanosensing controls the structure behind the architecture in plants. Current Opinion in Plant Biology, 2013. DOI : 10.1016/j.pbi.2013.06.006. [Publication scientifique]
[6] Landrein B., et al., How mechanical stress controls microtubule behavior and morphogenesis in plants: History, experiments and revisited theories. Plant Journal, 2013. DOI : 10.1111/tpj.12188. [Review]
[7] Burk D.H., et al., A katanin-like protein regulates normal cell wall biosynthesis and cell elongation. Plant Cell, 2001. DOI : 10.1105/tpc.13.4.807. [Publication scientifique]
[8] Burian A., et al., A correlative microscopy approach relates microtubule behaviour, local organ geometry, and cell growth at the Arabidopsis shoot apical meristem. Journal of Experimental Botany, 2013. DOI : 10.1093/jxb/ert352. [Publication scientifique]
[9] Aegerter-Wilmsen T., et al., Model for the regulation of size in the wing imaginal disc of Drosophila. Mechanisms of Development, 2007. DOI : 10.1016/j.mod.2006.12.005. [Publication scientifique]
[10] Schluck T., et al., Mechanical control of organ size in the development of the Drosophila wing disc. PLoS ONE, 2013. DOI : 10.1371/journal.pone.0076171. [Publication scientifique]
[11] Hervieux N., et al., A mechanical feedback restricts sepal growth and shape in Arabidopsis. Current Biology, 2016. DOI : 10.1016/j.cub.2016.03.004. [Publication scientifique]
[12] Hervieux N., et al., Mechanical shielding of rapidly growing cells buffers growth heterogeneity and contributes to organ shape reproducibility. Current Biology, 2017. DOI : 10.1016/j.cub.2017.10.033. [Publication scientifique]
[13] Oates A.C., What’s all the noise about developmental stochasticity? Development, 2011. DOI : 10.1242/dev.059923. [Review]
[14] Irvine K.D., et al., Mechanical control of growth: ideas, facts and challenges. Development, 2017. DOI : 10.1242/dev.151902. [Review]
Publié le 20/10/2024
Hervé Rabillé/Papier-Mâché/CC BY-NC-SA 4.0 2024Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.