Chargement de l'article...
Écriture : Bastien Malbert
Relecture scientifique : Anne-Sophie Masson
Relecture de forme : Éléa Héberlé et Line Kong A Siou
Temps de lecture : environ 11 minutes.
Thématiques : Biologie végétale (Biologie)
Publication originale : Block A., et al., The Pseudomonas syringae type III effector HopG1 targets mitochondria, alters plant development, and suppresses plant innate immunity. Cellular Microbiology, 2010. DOI : 10.1111/j.1462-5822.2009.01396.x

Dans une publication de 2010, une équipe de chercheurs de l’Université du Nebraska a montré qu’une protéine de la bactérie Pseudomonas syringae bloque la mise en place des défenses de la plante infectée. Les auteurs ont démontré que cette protéine, injectée dans la plante par la bactérie, est localisée dans les mitochondries, sortes de centrales énergétiques des cellules eucaryotes aussi impliquées dans la défense cellulaire. À l’image de la course aux armements lors de la guerre froide, la coévolution des bactéries pathogènes et des plantes leur a permis d’obtenir un véritable arsenal biologique, pouvant faire basculer le cours de chaque bataille.
À l’image de la course aux armements de la Guerre Froide, pendant laquelle soviétiques et américains investissent massivement dans la recherche et le développement de nouvelles technologies militaires en prévision d’un conflit frontal, les plantes et les bactéries ne se font aucun cadeau ! Ciblage des mécanismes de reconnaissance de l’ennemi, de communication entre les différents compartiments de la cellule, blocage des attaques par la plante, mise en place de défenses très spécifiques et ciblées… Interagissant depuis probablement plusieurs millions d’années, l’évolution des plantes et des bactéries ne s’est pas faite de façon indépendante. Dès lors que la bactérie évolue et gagne en virulence, c’est-à-dire en pouvoir pathogène, la plante subit une pression de sélection. Ici aussi, on peut parler de course aux armements afin de bloquer ou réussir l’infection.
Qu’est-ce qu’une infection réussie ?
Une infection correspond à la pénétration d’un microorganisme dans un être vivant, qui sera son hôte. En se multipliant, le microorganisme peut créer des lésions, altérant la santé de l’hôte : on parle alors de maladie. S’il est plutôt facile de définir ces termes, il est nettement plus compliqué de déterminer si une infection est réussie ou non. On peut supposer qu’à partir du moment où un microorganisme réussit à neutraliser les défenses et se multiplier chez son hôte, l’infection est considérée comme réussie.
La neutralisation des défenses des plantes passe par des attaques ciblées. C’est à ce moment qu’interviennent les protéines bactériennes appelées effecteurs. Présents chez la plupart des pathogènes, les effecteurs sont envoyés de la bactérie vers la cellule végétale lors de l’infection.
Cependant, toutes les bactéries connues pour pouvoir infecter les plantes sont-elles capables de réussir une infection sur tous les végétaux ? Même si l’attaque d’un arbre, ou d’un champ entier retient l’attention, l’infection est une rare exception puisque la majorité des interactions entre les plantes et les pathogènes ne donnent pas lieu à une attaque réussie [1].
Deux raisons expliquent cette incompatibilité. La première, mécanique, dépend de l’épaisseur de la paroi des cellules végétales. Plus large chez certaines espèces, les bactéries vont se retrouver littéralement prises de court pour déployer les effecteurs dans la cellule. La « seringue moléculaire » qui leur permet le déploiement d’effecteurs à travers la paroi peut parfois être trop petite pour perforer complètement ce mur protégeant les cellules des plantes. Quant à la seconde raison, elle dépend directement du système immunitaire des plantes.
Les plantes aussi ont un système immunitaire
Prenons l’exemple de Roger. Comme tout un chacun, ce grand gaillard de 17 ans possède un système immunitaire inné. Celui-ci voit arriver les pathogènes et met en place des réponses qui vont lui permettre de leur résister. Pour autant, malgré ses rencontres fortuites avec différents pathogènes, le système immunitaire de Roger ne s’en souvient pas. Il ne dispose pas d’un système immunitaire adaptatif. En plus de ça, l’immunité de Roger ne se fait pas par l’intermédiaire de cellules spécialisées — à l’image des globules blancs chez les humains. À l’inverse, chacune de ses cellules va apporter sa pierre à l’édifice immunitaire.
Est-ce que Roger souffre d’une maladie particulière ? Son système immunitaire est-il grandement fragilisé ? Eh bien non, Roger est en très bonne santé, et pour cause, Roger est un lierre (Figure 1).

À lui seul, Roger résume parfaitement les deux principales différences entre notre immunité et celle des plantes : pas de souvenir immunitaire de ses précédentes rencontres avec les pathogènes et pas de cellule spécifique impliquée uniquement dans la défense.
En regardant d’un peu plus près les réactions de Roger lorsqu’une bactérie pathogène se trouve dans son environnement proche, on se rend compte qu’il procède par étapes successives.
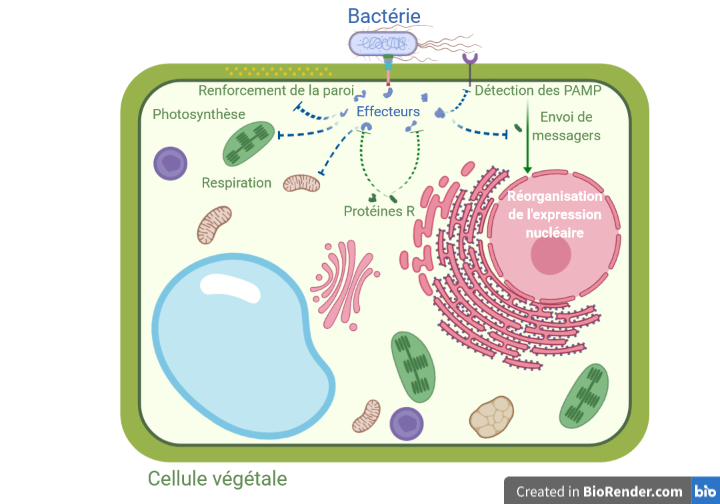
Roger va tenter de détecter des motifs moléculaires conservés, c’est-à-dire présents chez tous les pathogènes — PAMP de leur petit nom (pour Pathogen Associated Molecular Patterns). Ces PAMP peuvent être reconnus par des récepteurs situés à l’extérieur de la paroi des cellules de Roger. Suite à la détection de PAMP, des messages sont envoyés vers le noyau de la cellule qui contient la majeure partie de l’information génétique, où ils coordonnent les réponses de défense : renforcement de la paroi ou production de composés antimicrobiens par exemple. On parle alors d’immunité déclenchée par les PAMP.
En utilisant ses effecteurs, le pathogène peut procéder à différentes attaques ciblées à l’intérieur des cellules de Roger : blocage des messagers, du déploiement de molécules de défense, etc. Avec cette stratégie, les bactéries peuvent rapidement bloquer l’immunité déclenchée par les PAMP et tirer profit de la situation si la plante ne réagit pas.
Roger n’a toutefois pas pour autant dit son dernier mot. Sa dernière arme, et non des moindres, consiste à utiliser des protéines de résistance capables de reconnaître les effecteurs. Ces protéines de résistance activent alors l’immunité déclenchée par les effecteurs. S’il y a un temps pour la parole, Roger est déjà passé aux actes : la cellule infectée met alors en place la réponse hypersensible, une forme de suicide cellulaire [2]. Si Roger ne peut plus profiter de cette cellule, le pathogène non plus ! Dans ce cas de figure, la plante est dite résistante et ne tombe pas malade. Mais toutes les plantes n’ont pas nécessairement de protéine de résistance efficace pour chaque pathogène rencontré.
La Figure 2 résume les différentes interactions entre la bactérie et la cellule végétale.
Des échanges permanents entre les plantes et les pathogènes
Qu’est-ce qui détermine la neutralisation des défenses de la plante et la réussite de l’infection ? Si les effecteurs sont correctement déployés, leur réussite dépendra de plusieurs paramètres. Est-ce que les effecteurs ciblent bel et bien les mécanismes de cette plante en particulier ? Est-ce qu’ils ont une rapidité d’action suffisante par rapport à la mise en place des défenses ? La plante possède-t-elle des protéines de résistance pouvant les reconnaître ? Pouvant permettre l’infection comme l’annihiler en se faisant repérer, les effecteurs déterminent souvent l’issue du combat entre les plantes et les bactéries, d’où l’intérêt majeur de la compréhension de leurs modes d’action.
De très nombreux chercheurs ont travaillé sur le rôle des effecteurs bactériens pendant l’infection. Parmi eux, une équipe de l’Université du Nebraska a étudié en 2010 le rôle de l’effecteur HopG1, protéine de la bactérie Pseudomonas syringae pv. tomato. Après avoir mis en évidence que cette protéine est très présente chez la plupart des bactéries pathogènes pour les plantes, les auteurs ont cherché à comprendre son rôle lors de l’infection.
Dans ce but, ils ont principalement travaillé avec la plante modèle Arabidopsis thaliana, l’arabette des dames, une sorte de chou sauvage très utilisé pour son cycle de vie rapide et sa capacité d’autofécondation.
Un effet direct sur le système immunitaire des plantes
Les plantes ayant deux lignes de défense principale : l’immunité déclenchée par les PAMP et l’immunité déclenchée par les effecteurs, les chercheurs se sont demandés si HopG1 est capable de les perturber.
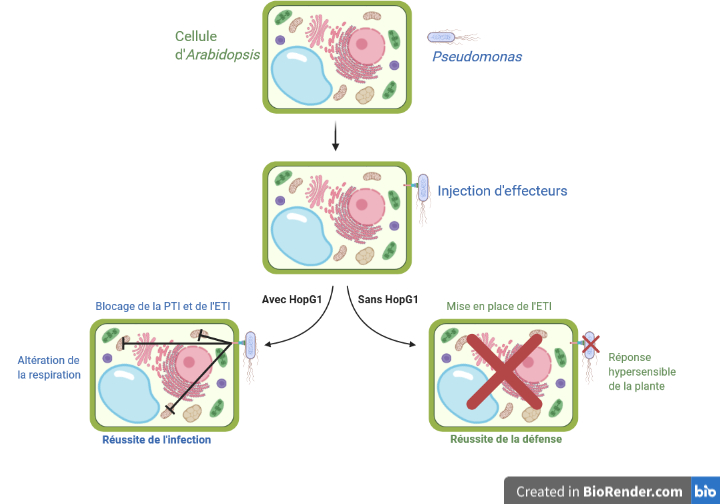
Pour cela, ils ont utilisé une approche assez contre-intuitive : étudier une souche de Pseudomonas incapable de produire l’effecteur HopG1. L’idée consiste à comparer une infection de plantes avec la bactérie sauvage — possédant l’effecteur HopG1 — à une infection de plantes avec la bactérie mutante qui ne l’a pas. Les auteurs ont alors remarqué que la réponse hypersensible est beaucoup plus importante avec la bactérie mutante. Ce suicide cellulaire correspond principalement à un mécanisme de l’immunité déclenchée par les effecteurs. Le fait que la réponse hypersensible soit plus importante dans le cas de l’infection par des bactéries sans HopG1 signifie cet effecteur peut bloquer cette immunité. Cela n’exclut pas pour autant un rôle dans un blocage éventuel de l’immunité déclenchée par les PAMP, les motifs moléculaires bactériens conservés.
Pour le vérifier, l’équipe de recherche a utilisé une autre stratégie. L’idée est de mettre au contact de la plante une souche bactérienne qui n’est pas capable d’envoyer ses effecteurs. Aucune chance alors de provoquer une immunité déclenchée par les effecteurs en absence d’effecteur ! Par contre, l’immunité déclenchée par les PAMP reste toujours possible puisque Pseudomonas va se trouver dans la plante, à l’extérieur des cellules mais à proximité de la paroi des cellules et de ses récepteurs. Pour cela, les chercheurs ont utilisé une souche mutée de la bactérie Pseudomonas rendue incapable d’envoyer HopG1 dans les cellules végétales. Ils ont ensuite infecté des lignées d’Arabidopsis thaliana. Étant donné que la bactérie ne peut pas injecter HopG1 à l’intérieur des cellules, mais que les chercheurs veulent étudier son rôle au cours de l’infection, ils ont génétiquement modifié les lignées d’Arabidopsis thaliana pour que ce soit elles qui expriment HopG1.
Les auteurs comparent alors les infections entre les plantes qui possèdent artificiellement HopG1 et les plantes témoins, c’est-à-dire les plantes non modifiées. Il s’avère que les bactéries se développent bien plus facilement dans les plantes exprimant HopG1 ! Cela signifie que HopG1 bloque aussi les défenses de la plante, indépendamment de l’immunité déclenchée par les effecteurs. HopG1 a donc aussi un rôle dans le blocage de l’immunité déclenchée par les PAMP.
Quel rôle pour HopG1 ?
Ces résultats montrent sans l’ombre d’un doute l’importance pour Pseudomonas d’avoir HopG1 à disposition lors de l’infection de l’arabette. On connaît dorénavant son implication dans le blocage de différents mécanismes de défense de la plante. Pour autant, on ne sait toujours pas par quel(s) mécanisme(s) moléculaire(s) HopG1 intervient. Avec quelle molécule de la plante interagit- il ? Dans quel compartiment cellulaire est-il localisé ? Quel club de football supporte-t-il ? Autant de questions qui restent en suspens…
En pratique, il n’existe pas de stratégie classique, évidente et surtout généralisable pour trouver la ou les cibles d’un effecteur. Il est donc extrêmement rare de la ou les identifier.
Chaque cellule de plante est délimitée par une paroi et une membrane. Mais elle contient elle-même différents compartiments. On peut citer le noyau dont nous avons déjà parlé, impliqué dans le maintien et l’expression des gènes, les chloroplastes où a lieu la photosynthèse, la vacuole où l’eau est stockée, etc. Afin de limiter le champ des possibles, les auteurs ont commencé par chercher le compartiment cellulaire de la plante dans lequel HopG1 se trouve au cours de l’infection. Pour cela, ils ont utilisé une approche très classique en biologie : ils ont fusionné HopG1 avec une protéine fluorescente verte, appelée GFP (pour Green Fluorescent Protein). Les chercheurs ont ensuite observé les feuilles au microscope, à la recherche de la fluorescence verte (Figure 3).
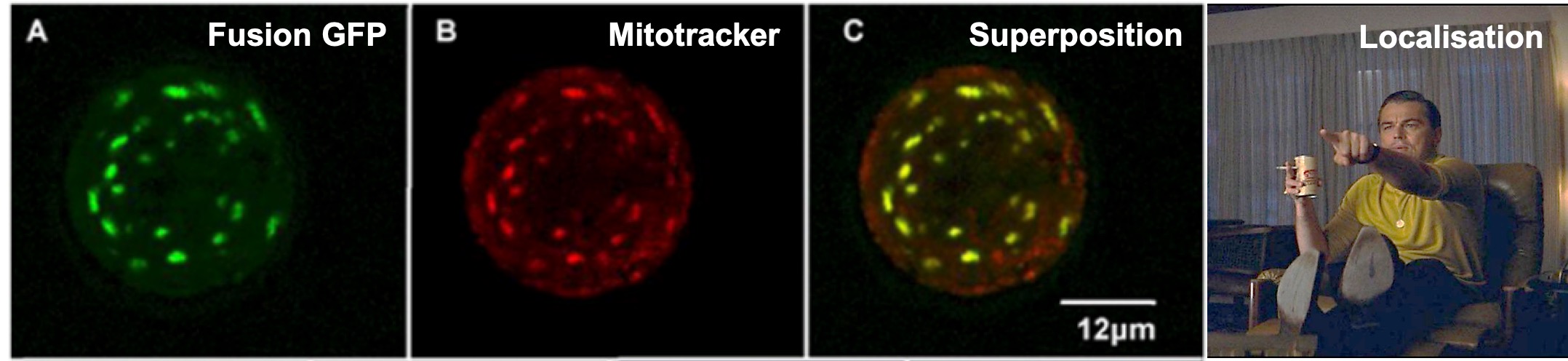
Ils ont constaté un résultat ressemblant à celui présenté Figure 3 : la fluorescence est localisée dans la mitochondrie. Indispensable à la grande majorité des cellules, la mitochondrie est le siège de la respiration cellulaire. En utilisant principalement du glucose et du dioxygène, la mitochondrie produit l’ATP, molécule énergétique indispensable à la réalisation de nombreuses réactions chimiques dans la cellule. Afin de voir si la respiration cellulaire était affectée par la présence de HopG1, les auteurs ont mesuré la quantité de dioxygène consommé par la plante. La comparaison d’une lignée d’Arabidopsis thaliana exprimant artificiellement HopG1 et d’une lignée témoin a alors mis en évidence une diminution de la respiration cellulaire en présence de HopG1.
Pour résumer, le travail de l’équipe des chercheurs a permis de démontrer l’importance de l’effecteur HopG1 dans le blocage des immunités déclenchées par les effecteurs et par les PAMP. Cette étude a également montré que HopG1 est localisé dans les mitochondries des cellules végétales, ainsi que son rôle dans l’altération de la respiration cellulaire (Figure Bilan).
Dans cette publication, les chercheurs de l’Université du Nebraska apportent non seulement beaucoup d’informations sur l’importance de HopG1 pour la réussite de l’infection de Pseudomonas, mais démontrent aussi pour la première fois qu’un effecteur bactérien peut cibler la mitochondrie. Leur hypothèse est que HopG1 interagirait avec une ou plusieurs protéines dans la mitochondrie, avec pour conséquence une suppression de la réponse hypersensible de la cellule, nécessaire à ce moment-là pour bloquer la progression du pathogène à l’échelle de la plante.
Ce travail de recherche montre également la difficulté de trouver la cible d’une protéine dans une cellule. De même, il n’est pas toujours facile de savoir si les conséquences que l’on observe sont un effet direct de l’interaction entre l’effecteur et sa cible, ou un effet collatéral sur un autre processus dans la cellule.
Mécanisme connu ou pas, Roger ne cache pas sa joie d’avoir tenu le choc. Mais il reste tout à fait conscient qu’un jour, un pathogène peut amener sa chute. « On ne vit qu’une fois ! Quand on va au front, on essaie de ne pas penser à ces choses là », conclut-il en agitant ses feuilles.
[1] Fu Z.Q. & Dong X., Systemic Acquired Resistance: Turning Local Infection into Global Defense. Annual Review of Plant Biology, 2013. DOI : 10.1146/annurev-arplant-042811-105606. [Publication scientifique]
[2] Jones J.D.G. & Dangl J.L., The plant immune system. Nature, 2006. DOI : 10.1038/nature05286. [Publication scientifique]
[3] Bentolila S., et al., RIP1, a member of an Arabidopsis protein family, interacts with the protein RARE1 and broadly affects RNA editing. PNAS, 2012. DOI : 10.1073/pnas.1121465109. [Publication scientifique]
Publié le 05/08/2020
 Bastien Malbert/Papier-Mâché/CC BY-NC-SA 4.0 2020
Bastien Malbert/Papier-Mâché/CC BY-NC-SA 4.0 2020Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.
Manuel d’infiltration des bactéries au pays des plantes
Écriture : Bastien Malbert
Relecture scientifique : Anne-Sophie Masson
Relecture de forme : Éléa Héberlé
Temps de lecture : environ 16 minutes.
Thématiques : Biologie végétale (Biologie)
Publication originale : Block A., et al., The Pseudomonas syringae type III effector HopG1 targets mitochondria, alters plant development, and suppresses plant innate immunity. Cellular Microbiology, 2010. DOI : 10.1111/j.1462-5822.2009.01396.x.
Dans une publication de 2010, une équipe de chercheurs de l’Université du Nebraska a montré qu’une protéine de la bactérie Pseudomonas syringae bloque la mise en place des défenses de la plante infectée. Les auteurs ont démontré que cette protéine, injectée dans la plante par la bactérie, est localisée dans les mitochondries, sortes de centrales énergétiques des cellules eucaryotes aussi impliquées dans la défense cellulaire. À l’image de la course aux armements lors de la guerre froide, la coévolution des bactéries pathogènes et des plantes leur a permis d’obtenir un véritable arsenal biologique, pouvant faire basculer le cours de chaque bataille.
Depuis la fin de la Seconde Guerre Mondiale, et ce jusqu’à la chute du bloc communiste en 1991, de fortes tensions opposent américains et soviétiques. D’abord idéologiques et politiques, les conflits se déplacent rapidement sur le terrain militaire, le plus souvent indirectement à travers des pays satellites comme lors de la guerre de Corée. En prévision d’un conflit plus frontal, les deux blocs investissent massivement dans la recherche et le développement de nouvelles technologies militaires : c’est la course aux armements.
Quel lien avec les bactéries infectant les plantes ? Leur stratégie est la même !
Ciblage des mécanismes de reconnaissance de l’ennemi, de communication entre les différents compartiments de la cellule, blocage des attaques par la plante, mise en place de défenses très spécifiques et ciblées, les plantes et les bactéries ne se font aucun cadeau. Interagissant depuis probablement plusieurs millions d’années, l’évolution des plantes et des bactéries ne s’est pas faite de façon indépendante. Dès lors que la bactérie évolue pour gagner en virulence, la plante subit une pression de sélection. On peut donc parler ici aussi de course aux armements afin de bloquer ou réussir l’infection.
Qu’est-ce qu’une infection réussie ?
Une infection correspond à la pénétration d’un microorganisme dans un être vivant, qui sera son hôte. En se multipliant, le microorganisme peut créer des lésions, altérant la santé de l’hôte : on parle alors de maladie. S’il est plutôt facile de définir ces termes, il est nettement plus compliqué de déterminer si une infection est réussie ou non. On peut supposer qu’à partir du moment où un microorganisme réussit à neutraliser les défenses et se multiplier chez son hôte, l’infection est considérée comme réussie.
La neutralisation des défenses des plantes passe par des attaques ciblées. C’est à ce moment qu’interviennent les protéines bactériennes appelées effecteurs. Encodés par le génome de la plupart des pathogènes, les effecteurs sont envoyées dans la cellule végétale lors de l’infection. Les plus connus et les mieux étudiés à ce jour sont les effecteurs de type III (ou T3E, pour Type III Effector) [*]. Le nom des T3E provient de la structure qui permet leur passage depuis la bactérie pathogène vers l’intérieur de la cellule végétale : le système de sécrétion de type III (ou T3SS, pour Type III Secretion System). Souvent vue comme une seringue moléculaire, cette structure permet aux bactéries de perforer la paroi des cellules végétales et d’y injecter les effecteurs. Cependant, toutes les bactéries phytopathogènes, c’est-à-dire connues pour pouvoir infecter les plantes, sont-elles capables de réussir une infection sur tous les végétaux ?
Même si l’attaque d’un arbre, ou d’un champ entier retient l’attention, l’infection est une rare exception puisque la majorité des interactions entre les plantes et les pathogènes ne donnent pas lieu à une attaque réussie [1].
Deux raisons expliquent cette incompatibilité. La première, mécanique, dépend de l’épaisseur de la paroi végétale. Plus large chez certaines espèces, les T3SS vont alors se retrouver littéralement pris de court pour déployer les effecteurs dans la cellule, la seringue moléculaire pouvant être trop petite pour perforer complètement ce mur qui protège les cellules des plantes. Quant à la seconde raison, elle dépend directement du système immunitaire des plantes.
Les plantes aussi ont un système immunitaire
Prenons l’exemple de Roger. Comme tout un chacun, ce grand gaillard de 17 ans possède un système immunitaire inné. Celui-ci voit arriver les pathogènes et met en place des réponses plus ou moins spécifiques qui vont lui permettre de leur résister. Pour autant, malgré ses rencontres fortuites avec différents pathogènes, Roger ne dispose pas de système immunitaire adaptatif clairement défini. En plus de ça, l’immunité de Roger ne se fait pas par l’intermédiaire de cellules spécialisées — à l’image des globules blancs chez les humains. À l’inverse, chacune de ses cellules va apporter sa pierre à l’édifice immunitaire.
Est-ce que Roger souffre d’une maladie particulière ? Son système immunitaire est-il grandement fragilisé ? Eh bien non, Roger est en très bonne santé, et pour cause, Roger est un lierre (Figure 1).
À lui seul, Roger résume parfaitement les deux principales différences entre notre immunité et celle des plantes : pas de système immunitaire adaptatif défini et pas de cellule immunitaire spécifique.
En regardant d’un peu plus près les réactions de Roger lorsqu’une bactérie pathogène se trouve dans son environnement proche, les chercheurs se sont rendu compte qu’il procède par étapes successives.
Roger va tenter de détecter des motifs moléculaires conservés chez les pathogènes (ou PAMP, pour Pathogen Associated Molecular Patterns). On peut citer l’exemple de la flagelline, une protéine très conservée du flagelle bactérien. Elle peut être reconnue par des récepteurs situés à l’extérieur de la paroi des cellules de Roger. La plupart des bactéries pathogènes vont entrer dans les plantes par des ouvertures naturelles ou des blessures. Une fois à l’intérieur de la plante, ces bactéries vont se loger dans l’apoplaste, l’espace situé entre les cellules, d’où l’importance pour les cellules végétales de posséder des récepteurs localisés sur l’extérieur de leur paroi. Suite à la détection de PAMP, des messages sont envoyés vers le noyau de la cellule où ils coordonnent les réponses de défense : renforcement de la paroi ou production de composés antimicrobiens par exemple. On parle alors d’immunité déclenchée par les PAMP.
En utilisant ses effecteurs, le pathogène peut procéder à différentes attaques ciblées dans les cellules de Roger : interception des messagers, inhibition du déploiement de molécules de défense, etc. Avec cette stratégie, les bactéries peuvent rapidement bloquer l’immunité déclenchée par les PAMP et tirer profit de la situation si la plante ne réagit pas.
Roger n’a toutefois pas pour autant dit son dernier mot. Sa dernière arme, et non des moindres, consiste à utiliser ses protéines R (pour Resistance) [**]. Capables de reconnaître les effecteurs — ou à défaut, leur présence suite à leur action sur une cible —, ces protéines vont activer le dernier volet de la défense des plantes, l’immunité déclenchée par les effecteurs. S’il y a un temps pour la parole, Roger est déjà passé aux actes : il met en place la réponse hypersensible, une forme de mort cellulaire programmée [2]. Si Roger ne peut plus profiter de cette cellule, le pathogène non plus ! Dans ce cas de figure, la plante est dite résistante et ne tombe pas malade. À l’inverse, si un pathogène est capable d’infecter une plante, et que celle-ci ne dispose pas de protéines R permettant de détecter les effecteurs déployés, la réponse hypersensible ne sera pas déployée. La Figure 2 résume les différentes interactions entre la bactérie et la cellule végétale.
Des échanges permanents entre les plantes et les pathogènes
Qu’est-ce qui détermine la neutralisation des défenses de la plante et la réussite de l’infection ? Si les effecteurs bactériens sont correctement déployés, leur réussite dépendra de la présence de leurs cibles chez la plante, de leur rapidité d’action par rapport à la mise en place des défenses ou encore de la présence de protéines R pouvant ou non les reconnaître. Pouvant permettre l’infection comme l’annihiler en se faisant repérer, les effecteurs déterminent souvent l’issue du combat entre les plantes et les bactéries, d’où l’intérêt majeur de la compréhension de leurs modes d’action.
De très nombreux chercheurs ont travaillé sur le rôle des effecteurs bactériens pendant l’infection. Parmi eux, une équipe de l’Université du Nebraska a étudié en 2010 le rôle de l’effecteur HopG1, protéine de la bactérie Pseudomonas syringae pv. tomato. Après avoir mis en évidence que cette protéine est très conservée chez les bactéries phytopathogènes, les auteurs ont cherché à étudier l’injection de HopG1 dans les cellules végétales par le système de sécrétion de type III (T3SS), sa localisation dans les cellules végétales et son rôle lors de l’infection. Dans ce but, ils ont principalement travaillé avec deux plantes modèles, très utilisées en biologie végétale. La première, Nicotiana benthamiana, est une plante proche du tabac appréciée pour ses grandes feuilles faciles à infiltrer par des bactéries, l’idéal pour simuler une interaction plante-bactérie. La seconde plante est Arabidopsis thaliana, l’arabette des dames, une plante de la même famille que les choux. Celle-ci est très utilisée pour son cycle de vie rapide, sa capacité d’autofécondation ou encore son génome de taille restreinte. Enfin, la plupart des méthodes utilisées sont en lien avec une stratégie très répandue en biologie végétale : la génétique inverse. Au lieu de partir d’un phénotype particulier pour en trouver le ou les gènes responsables, on crée artificiellement une ou plusieurs mutations affectant le gène que l’on souhaite étudier et on analyse le ou les phénotypes obtenus.
HopG1, un effecteur dans le vent les cellules végétales
Afin de vérifier si HopG1 est bien envoyé dans la plante par la bactérie lors de l’infection, les auteurs ont utilisé une stratégie portant un nom complexe, mais somme toute assez simple : la fusion adénylate cyclase. L’idée consiste à fusionner le gène codant pour la protéine HopG1 dans Pseudomonas avec le gène codant pour une enzyme qui s’appelle l’adénylate cyclase (abrégée en CyaA) (Figure 3). Cette enzyme est connue pour catalyser la réaction de conversion de l’Adénosine TriPhosphate (ATP), molécule énergétique universelle nécessaire à de nombreuses réactions chimiques, en Adénosine MonoPhosphate cyclique (AMPc), molécule un peu moins courante mais facile à quantifier. Si la fusion HopG1-CyA fonctionne et que la bactérie injecte bien HopG1 dans la cellule de la plante, on s’attend alors à observer une plus grande quantité d’AMPc dans la cellule. C’est exactement ce que les auteurs ont observé, mettant ainsi en évidence l’injection de HopG1 dans les cellules de tabac.
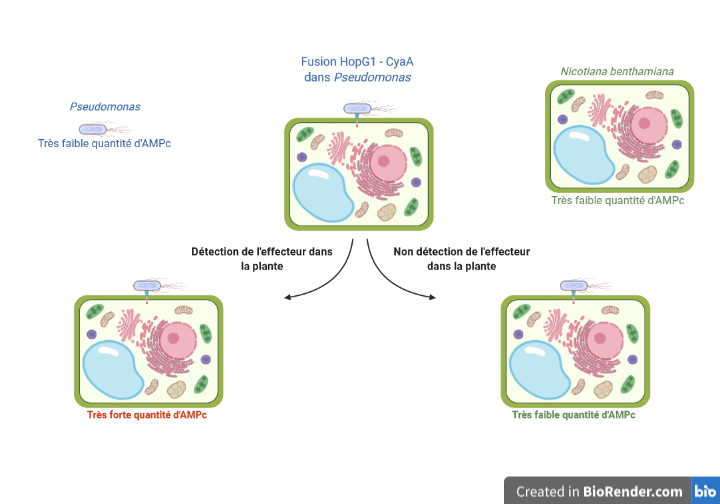
Mais pourquoi s’arrêter là alors que la génétique inverse nous permet d’en apprendre encore plus sur HopG1 ? Les chercheurs ont alors voulu étudier si HopG1 entre dans les cellules végétales à l’aide du T3SS, la seringue moléculaire permettant le passage des protéines bactériennes vers la cellule. Pour cela, ils ont à nouveau effectué une fusion HopG1-CyaA. Mais au lieu de travailler avec la souche sauvage de la bactérie Pseudomonas, celle trouvée dans la nature, l’équipe a travaillé sur une souche contenant une mutation pour un gène particulier, hrcC, qui code pour une protéine majeure du T3SS. Aussi, une fois mutée, la bactérie n’est plus capable de déployer son T3SS. En utilisant cette souche et la fusion HopG1-CyaA, aucune augmentation d’AMPc n’est observée dans les cellules végétales. Non seulement HopG1 est bien envoyé dans les cellules végétales en condition d’infection, mais en plus, on sait maintenant qu’il passe bien par le T3SS et non par un autre mécanisme.
Malgré cette belle découverte, le plus dur reste à venir pour l’équipe de recherche : trouver la cible de HopG1.
Un effet direct sur le système immunitaire des plantes
Afin de réduire les cibles possibles, plusieurs approches sont nécessaires. La première consiste à comprendre le rôle que joue HopG1 lors de l’infection. C’est dans cette optique que les auteurs ont exprimé le gène codant pour HopG1 directement dans la plante Arabidopsis thaliana. Le résultat n’a pas dû les décevoir, puisque la plante présente un phénotype assez extrême : infertilité, nanisme et surdéveloppement de très nombreuses feuilles (Figure 4).

Assez étonnamment, ce phénotype ne ressemble en rien à une plante infectée par Pseudomonas syringae. Comment interpréter ce résultat ? Plusieurs explications sont possibles selon les auteurs de la publication. La première serait que l’expression de HopG1 directement dans toutes les cellules de la plante l’amène à interagir avec des cibles qui lui sont normalement inaccessibles lors de l’infection, d’où ce phénotype inattendu. La seconde explication, qui n’exclut pas la première, serait que la ou les cibles de HopG1 joueraient un rôle non seulement dans la défense de la plante, mais aussi dans son développement.
À ce stade, les chercheurs savent que HopG1 joue un rôle dans l’infection, mais sans pouvoir déterminer lequel. Est-il capable de perturber la mise en place de l’immunité déclenchée par les PAMP ? De l’immunité déclenchée par les effecteurs ? En utilisant à nouveau une approche de génétique inverse, l’équipe a utilisé une souche de Pseudomonas mutée pour le gène codant pour HopG1, n’exprimant pas cette protéine. En infectant les plantes avec cette souche mutée, les auteurs ont observé une réponse hypersensible bien plus importante, en comparaison avec une infection par la souche sauvage. Ce suicide cellulaire correspond principalement à un mécanisme de défense de l’immunité déclenchée par les effecteurs, c’est-à-dire à la reconnaissance d’un effecteur ou de son action par une protéine R de défense de la plante. HopG1 jouerait donc un rôle dans le blocage ce type d’immunité, sans que cela n’exclut pour autant un rôle dans un blocage éventuel de l’immunité déclenchée par les PAMP.
Pour le vérifier, la souche de Pseudomonas mutée pour le gène hrcC incapable de déployer un T3SS fonctionnel est à nouveau utilisée pour infecter cette fois-ci les lignées d’Arabidopsis thaliana exprimant directement HopG1. L’idée des chercheurs est de mettre au contact de la plante une souche bactérienne qui n’est pas capable d’envoyer ses effecteurs à l’intérieur des cellules pour bloquer la défense de la plante. Aucune chance alors de déclencher une immunité déclenchée par les effecteurs en absence d’effecteur ! Pour autant, l’immunité déclenchée par les PAMP reste toujours possible puisque Pseudomonas va toujours se trouver dans l’apoplaste, à proximité de la paroi végétale et de ses récepteurs. Les auteurs comparent alors les infections entre deux lignées de plantes, l’une exprimant HopG1, l’autre non.
Il s’avère que les bactéries se développent bien plus facilement dans les plantes exprimant HopG1, même en absence d’immunité déclenchée par les effecteurs ! Cela suggère HopG1 joue également un rôle dans le blocage de l’immunité déclenchée par les PAMP.
Quel rôle pour HopG1 ?
Ces derniers résultats montrent sans l’ombre d’un doute l’importance pour la bactérie Pseudomonas d’avoir HopG1 à disposition lors de l’infection. On connaît dorénavant son implication dans le blocage de différents mécanismes de défense de la plante. Pour autant, on ne sait toujours pas par quel(s) mécanisme(s) moléculaire(s) HopG1 intervient. Avec quelle molécule de la plante interagit- il ? Dans quel compartiment cellulaire est-il localisé ? Quel club de football supporte-t-il ? Autant de questions qui restent en suspens…
En pratique, il n’existe pas de stratégie classique, évidente et surtout généralisable pour trouver la ou les cibles d’un effecteur. Il est donc extrêmement rare de la ou les identifier. Afin de limiter le champ des possibles, les auteurs ont néanmoins cherché le compartiment cellulaire de la plante dans lequel HopG1 se trouve pendant l’infection. Pour cela, ils ont utilisé une approche proche de celle utilisée pour démontrer son injection : ils ont fusionné HopG1 avec une protéine fluorescente, la GFP (pour Green Fluorescent Protein). En exprimant dans la plante la protéine fusion HopG1-GFP, les chercheurs ont ensuite observé les feuilles au microscope, à la recherche de fluorescence verte [***].
Les chercheurs ont constaté un résultat ressemblant à celui présenté Figure 5. Cela indique que la fluorescence est localisée dans un compartiment cellulaire important à la fois pour le développement et la réponse aux pathogènes pour les plantes : la mitochondrie.
Indispensable à la majorité des cellules, cette structure est le siège de la respiration cellulaire. En utilisant principalement du glucose et du dioxygène, la mitochondrie produit de l’ATP, la principale molécule énergétique des cellules. Afin de voir si la respiration cellulaire est affectée en présence de HopG1, les auteurs ont alors mesuré la quantité de dioxygène consommé par la plante à l’obscurité. Pourquoi faire l’expérimentation dans le noir alors que les plantes apprécient le feu des projecteurs ? Tout simplement pour que la photosynthèse et sa production de dioxygène ne viennent pas fausser les mesures. La comparaison d’une plante exprimant HopG1 à une plante sauvage met en évidence une diminution de la respiration cellulaire, avec une consommation de dioxygène réduite d’environ la moitié en présence de HopG1, montrant ainsi que la présence d’HopG1 entraîne une diminution de la respiration cellulaire.
Pour résumer, le travail de l’équipe de chercheurs a mis en évidence l’injection de HopG1 dans les cellules végétales lors de l’infection par Pseudomonas, son importance dans le blocage des immunités déclenchées par les effecteurs et par les PAMP dans la diminution de la respiration et enfin sa localisation mitochondriale (Figure Bilan).
Dans cette publication, les chercheurs de l’Université du Nebraska apportent non seulement beaucoup d’informations sur l’importance de HopG1 pour la réussite de l’infection de Pseudomonas, mais démontrent aussi pour la première fois qu’un effecteur bactérien peut cibler la mitochondrie. Leur hypothèse est que HopG1 interagirait avec une ou plusieurs protéines cibles dans la mitochondrie, avec pour conséquence une suppression de la réponse hypersensible de la cellule nécessaire à ce moment-là pour bloquer la progression du pathogène à l’échelle de la plante.
Non seulement ce travail de recherche démontre la difficulté de trouver la cible d’une protéine, mais aussi qu’il n’est pas toujours facile de savoir si les conséquences que l’on observe sont l’effet direct de l’interaction entre l’effecteur et sa cible, ou un effet collatéral sur un autre processus dans la cellule. En effet, chaque mécanisme perturbé et chaque conséquence de l’action de l’effecteur peut très bien être directe, par une interaction avec une ou des protéines par exemple, mais aussi une conséquence plus indirecte, lointaine. L’accumulation de signaux, parfois contradictoires, peut brouiller et largement complexifier la recherche d’une cible.
D’autres approches, comme la méthode TAP (pour Tandem Affinity Purification, purification par affinité en tandem), permettent d’aller à la pêche aux protéines d’intérêt dans les cellules, afin de déterminer avec quelles molécules elles interagissent. Plus complexes à mettre en place, ces techniques peuvent aboutir, en se basant sur les connaissances des autres protéines en interaction, à la détermination de l’action directe d’un effecteur dans une cellule.
Mécanisme connu ou pas, Roger ne cache pas sa joie d’avoir tenu le choc. Mais il reste tout à fait conscient qu’un jour, un pathogène peut amener sa chute. « On ne vit qu’une fois ! Quand on va au front, on essaie de ne pas penser à ces choses là », conclut-il en agitant ses feuilles.
[*] Un effecteur de Type III est une protéine injectée dans une cellule, animale ou végétale, par un pathogène utilisant un système de sécrétion. On parle d’effecteur de Type III lorsque celui-ci passe de la bactérie à la cellule végétale par un système de sécrétion de Type III. Chez les bactéries, 6 types de systèmes de sécrétion ont été identifiés jusqu’à présent. Ils diffèrent de par leur structure et le nombre de protéines qui les composent.
[**] Les protéines R sont responsables de la détection des effecteurs dans la cellule végétale, entraînant une réponse immunitaire spécifique et forte. Ces protéines procurent un avantage extrêmement important aux plantes dans la lutte contre certains pathogènes. Si aucune protéine R n’est capable de détecter les effecteurs d’un pathogène, et que les défenses non spécifiques ne sont pas suffisantes, le pathogène pourra continuer son infection.
[***] Les chercheurs auraient aussi pu fusionner HopG1 et la GFP dans la bactérie, puis attendre son injection dans la plante pour suivre la localisation de la fluorescence. Seulement, la protéine GFP est trop globuleuse pour passer par la seringue moléculaire T3SS, ce qui les a contraint à trouver une méthode alternative.
[1] Fu Z.Q. & Dong X., Systemic Acquired Resistance: Turning Local Infection into Global Defense. Annual Review of Plant Biology, 2013. DOI : 10.1146/annurev-arplant-042811-105606. [Publication scientifique]
[2] Jones J.D.G. & Dangl J.L., The plant immune system. Nature, 2006. DOI : 10.1038/nature05286. [Publication scientifique]
[3] Su W., et al., Conserved Endoplasmic reticulum-associated degradation system to eliminate mutated receptor-like kinase in Arabidopsis. PNAS, 2010. DOI : 10.1073/pnas.1013251108. [Publication scientifique]
[4] Bentolila S., et al., RIP1, a member of an Arabidopsis protein family, interacts with the protein RARE1 and broadly affects RNA editing. PNAS, 2012. DOI : 10.1073/pnas.1121465109. [Publication scientifique]
Publié le 05/08/2020
Bastien Malbert/Papier-Mâché/CC BY-NC-SA 4.0 2020Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.