Chargement de l'article...
Écriture : Camille Lemonnier
Relecture scientifique : Candice Michelot
Relecture de forme : Hervé Rabillé et Éléonore Pérès
Temps de lecture : environ 12 minutes.
Thématiques : Comportement animal (Biologie), Écologie
Publication originale : Orgeret F., et al., Exploration during early life: distribution, habitat and orientation preferences in juvenile king penguins. Movement Ecology, 2019. DOI : 10.1186/s40462-019-0175-3.
Accès sur HAL (pdf).

Sans carte ni GPS, difficile de se repérer, surtout en territoire peu ou pas connu. Pourtant, beaucoup d’animaux sont capables de se déplacer et de se repérer sur des distances impressionnantes. L’exemple le plus parlant est celui des oiseaux migrateurs qui sont capables de rejoindre leur site d’hivernage depuis leur site de reproduction, parfois éloignés de plusieurs milliers de kilomètres, et ce, année après année. D’autres animaux parcourent de longues distances entre leurs sites de reproduction et de nourrissage et sont également capables de retrouver leur chemin malgré la distance. C’est le cas des manchots royaux. Mais comment ces animaux arrivent-ils à se repérer ?
Le cycle de vie des manchots : entre terre et mer
Les manchots sont des oiseaux marins plongeurs au cycle de vie contraignant, rythmé par des périodes de reproduction sur terre et des périodes d’alimentation en mer. Cette dualité les rend vulnérables aux conditions environnementales propres à ces deux milieux, et conditionne leur survie. Chaque adulte doit être efficace pour trouver sa nourriture et éviter les prédateurs en mer, afin d’assurer sa propre survie et se reproduire avec succès sur terre.
Les manchots royaux ont un cycle de vie qui se démarque par sa longueur : il faut plus d’un an pour qu’un couple d’adultes élève un petit (Figure 1), contrairement à d’autres espèces comme le manchot Adélie pour lequel le cycle s’étale sur environ 6 mois. Ces oiseaux vivent en colonie sur les îles subantarctiques (à la limite nord de l’océan Antarctique), où les adultes reproducteurs se rassemblent au début de l’été austral entre octobre et novembre pour la reproduction et l’élevage des poussins. Un œuf unique par couple est pondu fin novembre. Les deux partenaires d’un couple se relaient alors au nid pour le garder : pendant que l’un part en mer se nourrir, le second incube l’œuf sur ses pattes. L’éclosion a lieu 55 jours après la ponte, après laquelle les parents continuent d’alterner entre des périodes de jeûne à terre pour garder le poussin et des voyages en mer destinés à la restauration de leurs réserves alimentaires ainsi qu’à l’alimentation du poussin. Les poussins sont thermiquement indépendants un mois après l’éclosion. Ils forment alors des groupes avec d’autres, appelés crèches, pour se tenir chaud pendant que les parents partent simultanément en mer se nourrir et reviennent de moins en moins souvent à terre. À la fin de l’été austral (mi-avril), les poussins ayant acquis suffisamment de réserves débutent une période de jeûne hivernal qui s’étale jusqu’en septembre. Les parents partent en mer pour rejoindre la limite de la banquise antarctique située à plusieurs centaines de kilomètres, et ne reviennent à terre que rarement pendant cette période pour nourrir les poussins. Ceux-ci finissent leur croissance et quittent la colonie pour se nourrir par eux-mêmes en mer mi-novembre, après avoir mué. Les adultes parvenant à élever leur poussin jusqu’à terme (mi-novembre) ne prennent pas part à la reproduction l’année suivante, la saison de reproduction ayant déjà commencé au moment du départ du jeune poussin de l’année précédente.
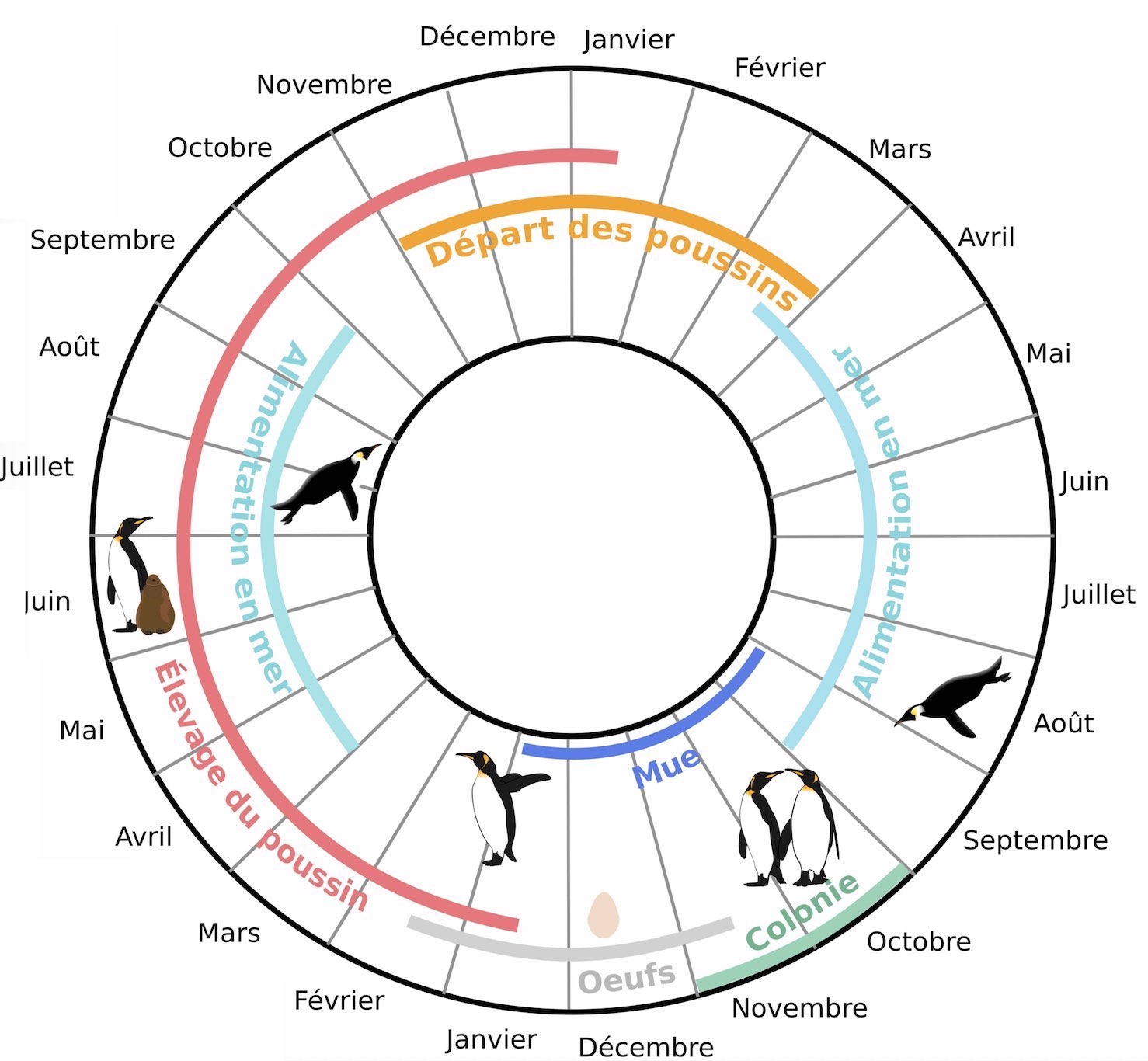
Ce cycle annuel hors normes est une des raisons qui poussent les chercheurs à tenter de mieux comprendre les comportements et les adaptations que ces oiseaux présentent face à leur environnement extrême et aux différentes contraintes auxquelles ils font face dans les îles subantarctiques. Ces contraintes peuvent être liées à l’environnement (la présence de prédateurs ou la proximité des autres adultes souvent agressifs dans la colonie, les conditions météorologiques extrêmes) ou d’ordre biologique (état de santé des adultes, conditions corporelles). En particulier, les périodes de jeûne et de mue sur terre sont très coûteuses en énergie pour les manchots. Pour restaurer leurs réserves corporelles (leur gras), les manchots doivent d’abord nager sur parfois plusieurs centaines de kilomètres pour rejoindre des zones où la nourriture est abondante. Ainsi, être capable de cibler efficacement des zones riches en proies est une qualité importante chez les manchots.
Les adultes ne prenant pas part à la reproduction ainsi que les jeunes manchots dont c’est le premier départ en mer (les juvéniles) s’engagent dans un voyage d’environ un an, pendant lequel ils parcourent plusieurs milliers de kilomètres pour se nourrir. Comment ces oiseaux arrivent-ils à se diriger vers des zones riches en ressources alimentaires dans un environnement aussi vaste que l’océan ? C’est la question que se sont posée les auteurs de la publication présentée ici.
L’orientation en mer : comment rejoindre efficacement de petites zones géographiques ?
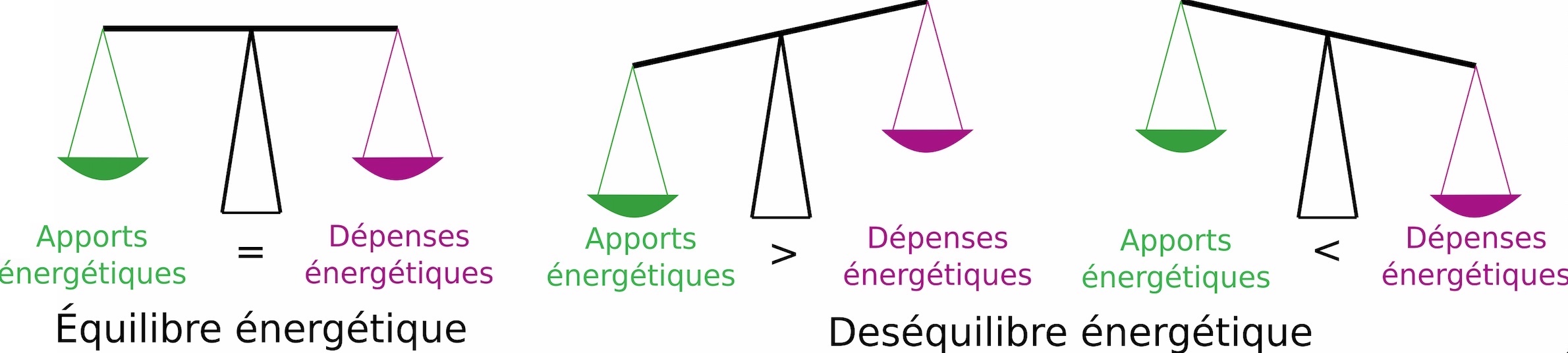
Les manchots doivent rejoindre rapidement des lieux riches en ressources alimentaires afin de ne pas dépenser plus d’énergie à la recherche de nourriture que ce qu’ils ne peuvent récupérer en mangeant (on parle de balance énergétique, Figure 2).
Les lieux présentant une forte abondance en proies se trouvent généralement au niveau de zones océaniques précises (comme les fronts océaniques) [**]. Ces zones riches en aliments ne sont pas fixes dans les océans, leur taille et leur emplacement varient selon les conditions environnementales (suivant les courants océaniques par exemple) et peuvent s’étendre sur plusieurs kilomètres. Une partie de l’article d’Orgeret et de ses collègues démontre notamment que ces mécanismes seraient en partie innés : les juvéniles partant en mer pour la première fois semblent capables de rejoindre ces zones sans apprentissage préalable auprès d’adultes plus âgés.
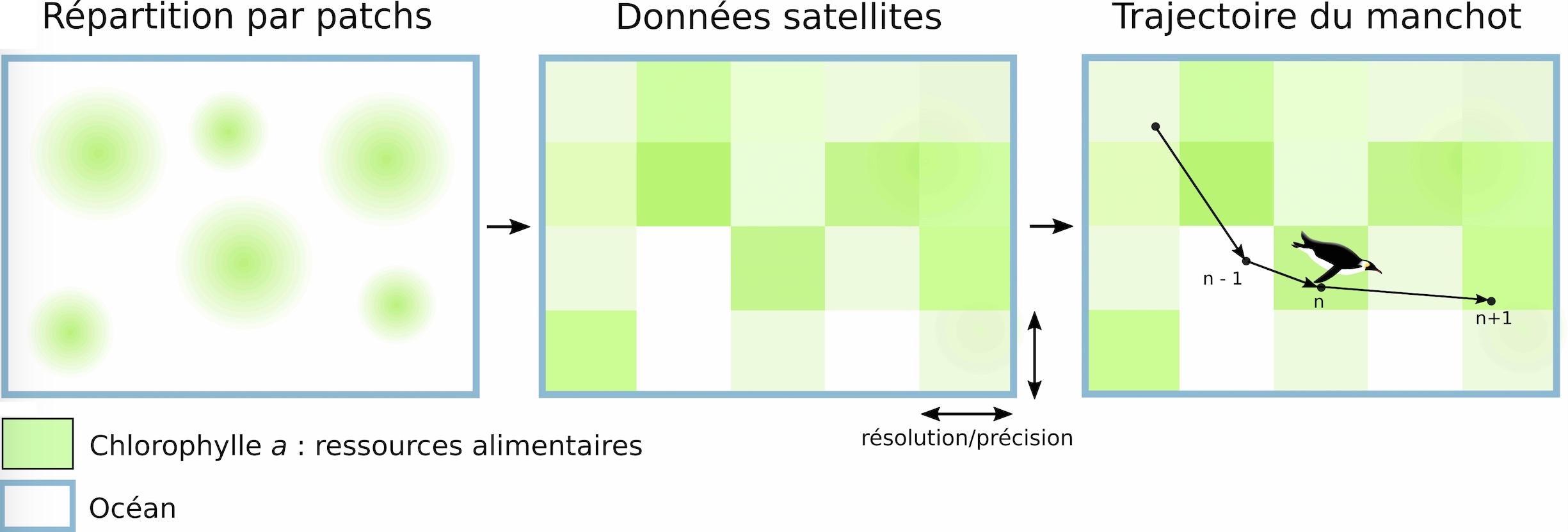
Si l’on zoome sur ces lieux de forte abondance en proies, on remarque que les ressources alimentaires n’y sont pas réparties de manière uniforme (c’est-à-dire avec la même concentration partout), mais elles sont plutôt distribuées en zones isolées, appelées patchs. Ce sont des zones qui se différencient par une concentration plus forte en ressources (ici, en proies) par rapport aux zones voisines (Figure 3). Les chercheurs ont identifié plusieurs moyens par lesquels les manchots peuvent identifier et cibler ces patchs : l’un de ces moyens est l’utilisation d’indices environnementaux, c’est-à-dire des paramètres physiques ou chimiques du milieu comme la vitesse et la force du vent ou du courant, ou la présence de certaines molécules par exemple.
Comprendre le comportement en mer des manchots : de la collecte des données à l’analyse
Les chercheurs se sont intéressés aux mouvements des manchots à fine échelle, c’est-à-dire sur des échelles de temps et de distance assez réduites par rapport au trajet complet des oiseaux, afin d’être en mesure de déceler d’éventuels liens entre les mouvements des manchots et des caractéristiques de leur environnement. Plus particulièrement, ils ont regardé l’orientation de leurs mouvements en fonction de différents paramètres environnementaux, afin de déterminer si certaines variations spatiales de ces paramètres peuvent influencer les déplacements des manchots.
Suivre les mouvements des manchots Les manchots sont suivis pendant leurs voyages en mer grâce à des balises Argos [***] fixées sur leurs plumes, qui permettent d’enregistrer leur position géographique à une fréquence régulière sans avoir à les observer directement. Grâce aux données de localisation des manchots, les chercheurs ont pu extraire les zones où les manchots se nourrissent.
Extraire les paramètres environnementaux correspondant Les chercheurs ont extrait tout au long des voyages des manchots différents paramètres environnementaux, comme la force et la direction des vents ou des courants, ainsi que la concentration en chlorophylle a, qui permet de représenter la concentration en ressources alimentaires disponibles [****]. Ces données sont récoltées grâce à des programmes d’observation de la météo ou des océans [*****].
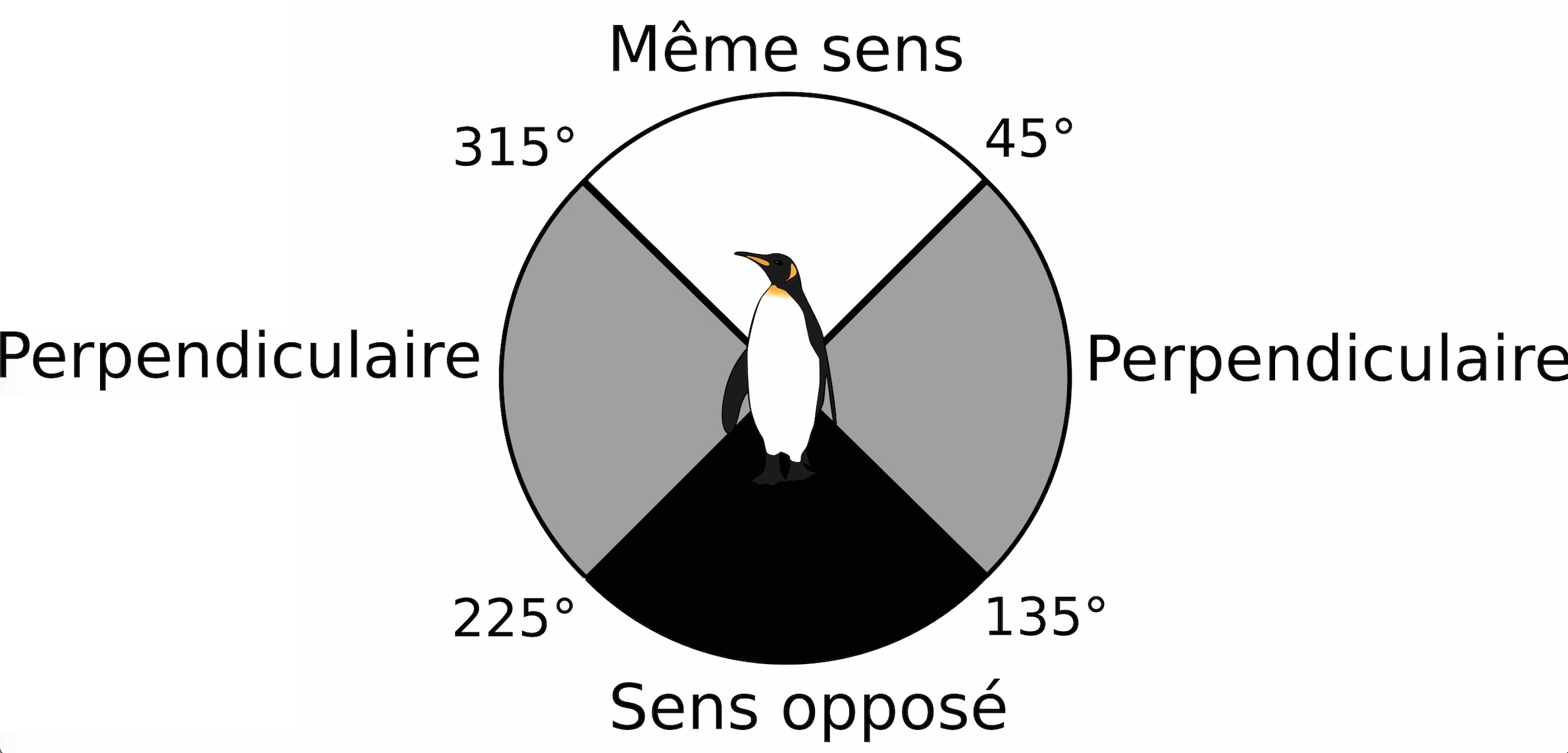
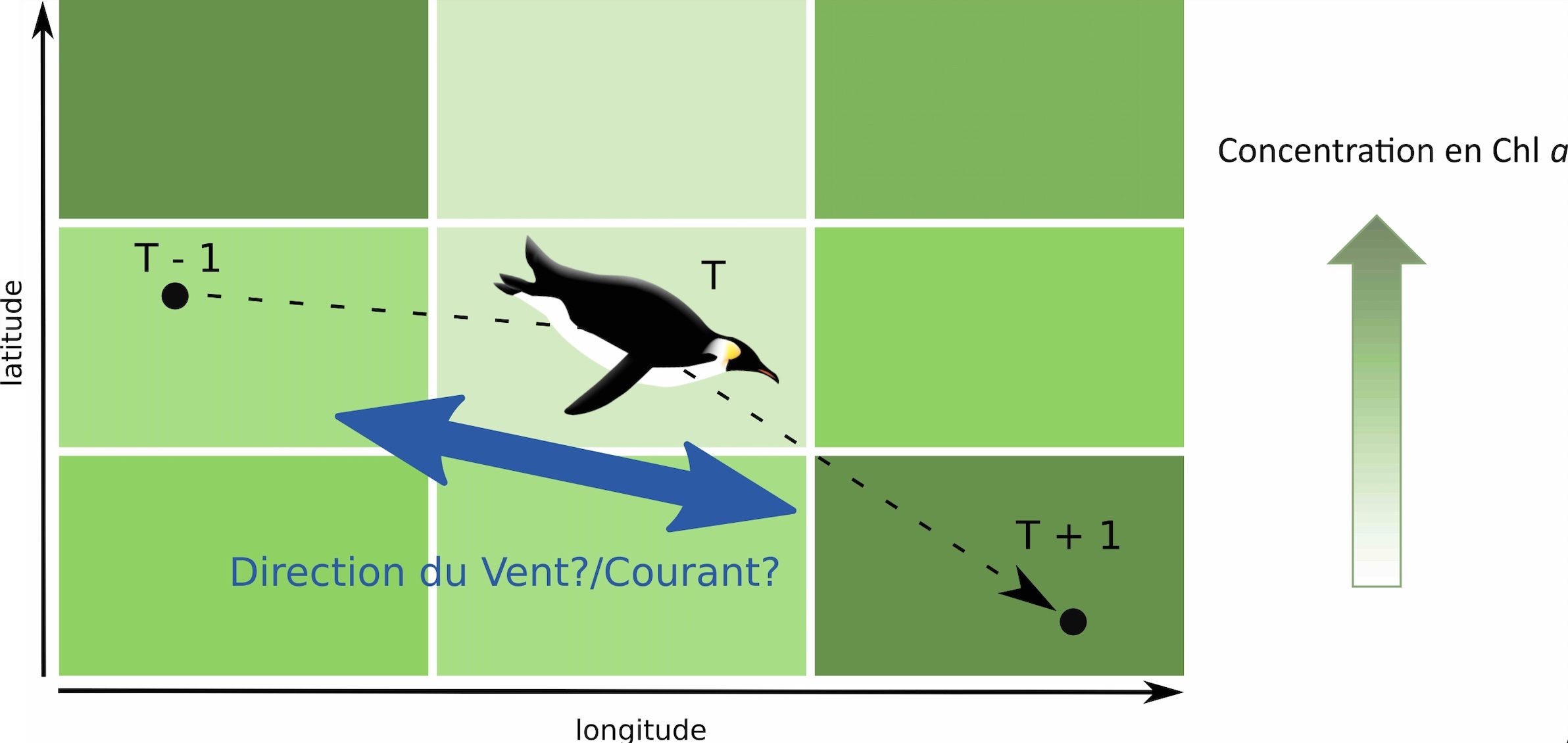
Comparer orientation des manchots et concentration en ressource Les chercheurs ont analysé les données selon un protocole bien particulier : pour chaque localisation d’un manchot (un point donné), on relève la concentration en chlorophylle a associée à la position précédente et celle à la position suivante, ce qui donne 3 positions. Cette première étape permet de déterminer s’il existe un changement progressif de la concentration de chlorophylle a sur la trajectoire de l’animal, autrement dit, si cette concentration varie entre ces 3 positions (augmente ou diminue, Figure 3). Ensuite, les chercheurs comparent l’orientation du mouvement du manchot (par exemple, s’il se dirige vers le Nord entre le point n-1 et n puis vers l’Est entre les points n et n+1) avec l’orientation du vent et du courant. On peut alors, pour chaque position de l’oiseau, calculer un angle entre son orientation et celle du vent/courant. Ces angles sont classés en 3 catégories : même sens, sens opposé ou sens perpendiculaire (Figure 4).
Finalement, les chercheurs comparent pour chaque position du manchot les catégories d’angles relevées (pour le vent et le courant) et les valeurs de concentration en chlorophylle a. Des modèles statistiques permettent de calculer la probabilité que chaque catégorie d’angle soit observée. Cette analyse permet de déterminer si, de manière générale, il existe un lien entre l’orientation des manchots, le sens du vent ou du courant et la présence de ressources alimentaires sur leur trajectoire.
Des manchots soumis aux paramètres environnementaux
La mise en œuvre du protocole précédent a permis d’obtenir les résultats illustrés dans la Figure 5. On remarque que lorsque la concentration en chlorophylle a augmente entre la position précédente et la suivante, la probabilité pour que les manchots s’orientent dans le sens opposé au vent augmente fortement et ce, aussi bien pour les juvéniles que pour les autres adultes non-reproducteurs (Figure 5A). En revanche, le courant (Figure 5B) ne semble pas avoir d’influence sur l’orientation des manchots par rapport à la concentration en chlorophylle a.
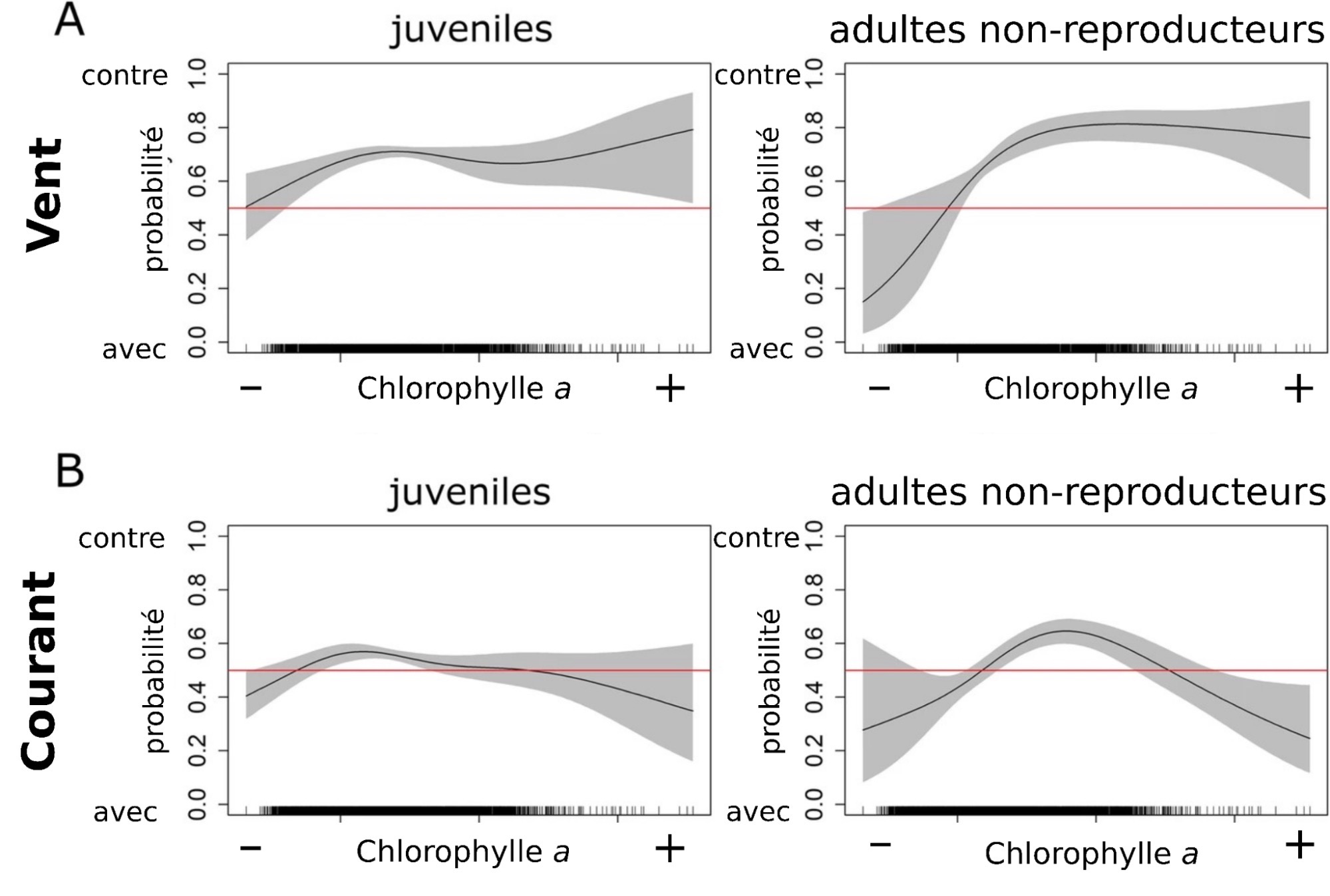
L’absence de relation claire entre le courant et l’orientation des manchots (Figure 5B, droite et gauche) peut facilement s’expliquer par l’absence de repère fixe permettant aux manchots de ressentir le courant. Ce repère fixe pourrait être le fond océanique par exemple, mais comme les manchots se déplacent en eau libre, ils ne peuvent pas vraiment profiter de ce repère. En revanche, il est plus probable que les manchots s’orientent par rapport au sens du vent (Figure 5A) car on observe bien une relation sur la figure, plus particulièrement dans le sens opposé au vent lorsque la concentration en chlorophylle a augmente le long de la trajectoire (vers le + sur la Figure 5). Autrement dit, si une zone riche en proie se trouve en amont du vent, il est probable que les manchots remontent contre le vent pour se diriger vers cette zone. Une hypothèse probable expliquant ce phénomène est que ces zones de forte productivité relâchent dans l’air des molécules qui sont ensuite transportées par le vent. Les manchots seraient capables de sentir ces molécules lorsqu’ils reviennent à la surface pour respirer, comme observé chez d’autres espèces (comme les tortues [1]). Cependant, on ne connaît pas exactement les capacités olfactives des manchots et de nouvelles études seraient nécessaires afin d’estimer dans quelles mesure les manchots pourraient utiliser des odeurs (des phéromones par exemple) portées par les vents pour s’orienter.
Conclusion
Cette étude fait donc le lien entre l’influence des paramètres environnementaux (notamment le sens du vent) et la distribution des ressources dans l’océan sur les mouvements des manchots. Cette étude permet également d’émettre des hypothèses quant au mécanisme permettant aux manchots de cibler ces zones de forte production alimentaire en évoquant la possibilité que les manchots détectent dans l’air des molécules transportées par le vent en provenance de ces zones.
L’étude de l’alimentation des manchots, et notamment les mécanismes et les adaptations à leur cycle de vie contraignant, permet de mieux comprendre les conséquences des modifications environnementales (au niveau des courants, des vents, de la disponibilité en proies dans l’océan, les variations climatiques sur terre) sur les populations de manchots. Ces études permettent aussi de mettre en place des solutions afin de préserver les zones importantes pour la survie des organismes qui vivent dans ces milieux et ainsi de sauvegarder ces écosystèmes.
[*] Photographie également publiée dans l’ouvrage Aventures Australes aux éditions Omniscience.
[**] Les fronts océaniques sont des zones où des masses d’eaux distinctes se rencontrent. Ces masses d’eau peuvent se différencier par plusieurs paramètres physiques ou chimiques : la direction des courants qui les entraînent, leur température, leur salinité ou d’autres encore. La plupart de ces fronts sont éphémères. En revanche, les fronts liés au courant circulaire qui entoure l’Antarctique durent souvent plus longtemps et sont à l’origine de bloom, c’est-à-dire d’une forte productivité organique synonyme de disponibilité en ressources alimentaires. La rencontre des masses d’eaux froides de l’océan Antarctique avec des masses d’eau plus chaudes des autres océans (ex : océan Indien) entraîne une augmentation de la biomasse (phytoplancton), c’est-à-dire de ressources alimentaires disponibles, dans ces zones. C’est un des lieux de nourrissage de prédilection pour les manchots pendant l’été (période pendant laquelle le front est actif).
[***] Le système ARGOS (Advanced Research and Global Observation Satellite) est un système satellitaire de localisation comparable au système GPS (Global Positioning System), mais il fonctionne différemment et ne donne pas les positions avec la même précision. Les balises ARGOS transmettent les données de localisation par voie satellitaire sans les stocker. Les données peuvent être alors téléchargées à distance, alors que les balises GPS collectent et stockent les données de localisation : il faut donc récupérer les balises pour avoir accès aux données. Pour plus d’informations sur les balises ARGOS et leur intérêt pour l’étude des manchots, vous pouvez consulter cette vidéo du CNES. Vous pouvez également lire ce papier mâché sur une autre utilisation possible du système ARGOS.
[****] La chlorophylle a est un élément indispensable pour la réalisation de la photosynthèse, réaction de base du monde végétal qui permet de convertir une partie de l’énergie solaire et de dioxyde de carbone (CO2) en matière organique. Dans les océans, c’est principalement le phytoplancton qui contient cette chlorophylle et réalise la photosynthèse. C’est ce que l’on appelle la production primaire, car le phytoplancton est au début des chaînes alimentaires. La concentration en chlorophylle a est un excellent paramètre permettant de connaître la productivité primaire d’un endroit et est souvent utilisée comme un indicateur de la disponibilité en ressources alimentaires pour d’autres espèces.
[*****] Les données proviennent du Global Forecast System of the United States National Weather Service et du Copernicus Marine Environment Monitoring Service of the European Union.
[1] Tortues : le parfum mortellement attractif du plastique marin. Sciences et Avenir, 2020. [Article de presse]
Publié le 19/10/2021
 Camille Lemonnier/Papier-Mâché/CC BY-NC-SA 4.0 2021
Camille Lemonnier/Papier-Mâché/CC BY-NC-SA 4.0 2021Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.
Écriture : Camille Lemonnier
Relecture scientifique : Candice Michelot
Relecture de forme : Hervé Rabillé et Éléonore Pérès
Temps de lecture : environ 19 minutes.
Thématiques : Comportement animal (Biologie), Écologie
Publication originale : Orgeret F., et al., Exploration during early life: distribution, habitat and orientation preferences in juvenile king penguins. Movement Ecology, 2019. DOI : 10.1186/s40462-019-0175-3.
Accès sur HAL (pdf).
Sans carte ni GPS, difficile de se repérer, surtout en territoire peu ou pas connu. Pourtant, beaucoup d’animaux sont capables de se déplacer et de se repérer sur des distances impressionnantes. L’exemple le plus parlant est celui des oiseaux migrateurs qui sont capables de rejoindre leur site d’hivernage depuis leur site de reproduction, parfois éloignés de plusieurs milliers de kilomètres, et ce, année après année. D’autres animaux parcourent de longues distances entre leurs sites de reproduction et de nourrissage et sont également capables de retrouver leur chemin malgré la distance. C’est le cas des manchots royaux. Mais comment ces animaux arrivent-ils à se repérer ?
Qu’est-ce que la navigation ?
Les animaux sont des êtres vivants motiles, c’est-à-dire capables de se mouvoir de façon spontanée et active (à ne pas confondre avec mobile : simple fait de pouvoir être déplacé). Cette capacité dépend de beaucoup de paramètres morphologiques et anatomiques, et repose aussi sur la consommation d’énergie. Pour décrire un mouvement, on s’intéresse à la trajectoire et à la vitesse de l’objet. Ainsi, décrire le mouvement d’un individu est relativement aisé. En revanche, comprendre comment et pourquoi les animaux prennent des décisions quant à leur trajectoire et leur vitesse lorsqu’ils sont en mouvement (ce qu’on appelle la navigation) est une tâche bien plus ardue. Plusieurs études ont démontré que les animaux pouvaient se servir d’indices présents dans leur environnement pour se diriger, comme par exemple des paramètres géomagnétiques, chimiques ou hydrodynamiques [1]. L’utilisation de ces indices et les décisions de navigation des animaux dépendent d’une part des échelles spatiales et temporelles auxquelles on se situe (courte ou longue distance, saisons, etc.), mais également de facteurs liés aux individus eux-mêmes (âge, sexe, condition corporelle). Cependant, une question demeure : comment les individus apprennent-ils à naviguer, et comment ces connaissances sont-elles transmises ?
Apprendre à naviguer : mimétisme, talent inné ou conditionnement par l’environnement ?
Plusieurs hypothèses ont été formulées pour tenter d’expliquer comment les animaux arrivent à s’orienter dans l’espace lors de longs trajets (en période ou en distance), et comment cette compétence est acquise et transmise à travers les générations :
- par apprentissage et/ou imitation d’autres individus de la même espèce plus expérimentés/âgés ;
- par l’acquisition de comportements de mouvement innées, acquis par la transmission de l’information génétique ;
- par l’utilisation des paramètres environnementaux (vent, odeur, courant, etc.).
À noter qu’aucune de ces hypothèses n’est exclusive et qu’il y a fort à parier que les animaux utilisent en réalité une combinaison de ces mécanismes.
Les manchots : des plongeurs et explorateurs hors pairs
Les manchots sont des oiseaux marins plongeurs. Pour cette espèce, être capable de se déplacer efficacement en mer et de se repérer est crucial pour la survie [2]. Les manchots sont des prédateurs marins qui se nourrissent de krill, poissons et céphalopodes [3] au large de leur colonie et peuvent se déplacer en mer sur de très longues distances [4]. Les conditions en mer sont souvent très éprouvantes : la plongée, la nage [5] et la thermorégulation [6] requièrent une grande dépense énergétique, et les manchots sont donc soumis à rude épreuve. Il est alors impératif de rejoindre rapidement les lieux d’alimentation et les lieux de reproduction une fois la bonne saison arrivée. De plus, les manchots sont très fidèles à leur site de reproduction [7], c’est-à-dire que les adultes sont capables de retourner vers leur colonie d’origine le moment venu. On peut donc se demander par quels mécanismes les manchots parviennent à réaliser ces mouvements saisonniers année après année et comment ces compétences sont transmises aux jeunes générations de manchots.
Dans la publication présentée ici, Orgeret et ses collègues s’intéressent à deux classes d’âge, appelées phases, peu connues du cycle de vie des manchots royaux (Aptenodytes patagonicus) dans une colonie située sur l’île de la Possession, dans l’archipel de Crozet (îles subantarctiques) : les juvéniles lors de leur premier voyage en mer (un an après leur éclosion), et les adultes ne prenant pas part à la reproduction : les adultes non-reproducteurs.
Les manchots royaux sont distribués de manière circumpolaire, et se reproduisent dans les îles subantarctiques. Cette espèce présente le cycle annuel (c’est-à-dire la façon dont la période de reproduction et de croissance des poussins en colonie sur terre et la période en mer s’enchaînent) le plus long de toutes les espèces de manchots, qui s’étend sur plus d’une année. Les adultes reproducteurs arrivent à la colonie entre les mois d’octobre et novembre. Un œuf unique est pondu fin novembre. Les deux partenaires d’un couple alternent alors la garde de l’œuf : pendant que l’un part en mer se nourrir, le second garde l’œuf sur ses pattes pour l’incuber. L’éclosion a lieu 55 jours après la ponte. Après l’éclosion, les parents continuent d’alterner entre des périodes de jeûne à terre pour garder leur poussin et des voyages en mer destinés à la restauration de leurs réserves alimentaires ainsi qu’à l’alimentation des petits. Un mois après l’éclosion, les poussins sont thermiquement autonomes et forment des crèches avec d’autres pour se tenir chaud, pendant que les parents partent simultanément en mer se nourrir, et reviennent moins régulièrement à terre. À la fin de l’été austral (mi-avril), les poussins ayant acquis suffisamment de réserves débutent une période de jeûne hivernal qui s’étalera jusqu’à l’hiver (septembre), pendant que les adultes partent en mer, parfois très loin, jusqu’à la limite de la banquise antarctique. Les parents reviennent à terre plus ou moins fréquemment pendant cette période pour assurer l’alimentation des poussins, qui finissent leur croissance et quittent la colonie pour se nourrir d’eux-mêmes en mer mi-novembre, après avoir mué. Les adultes commencent quant à eux une période de mue coûteuse en énergie, avant de se reproduire l’année suivante. Ainsi, les juvéniles de moins d’un an et les adultes s’étant reproduit avec succès lors d’un cycle donné ne feront pas face aux contraintes de reproduction au cycle suivant. Comparer le comportement en mer de ces deux catégories d’individus qui ne se reproduisent pas, lors du cycle suivant, permet alors de mieux comprendre les mécanismes par lesquels ils s’orientent et retrouvent leur chemin jusqu’à la colonie.
Suivre le mouvement des manchots royaux
Les longues périodes que passent les manchots royaux en mer rendent leur étude très difficile. En effet, ces animaux peuvent séjourner plus d’un an en mer et se dispersent sur de vastes étendues. De plus, les manchots sont très souvent invisibles sous la surface de l’eau, les observations directes (depuis des bateaux) sont donc insuffisantes pour étudier leur mouvement. Ainsi, les chercheurs doivent se baser sur des observations indirectes. Grâce aux nouvelles techniques de télémétrie, les auteurs de l’étude présentée ici sont parvenus à suivre le mouvement de manchots royaux pendant la totalité de leur voyage en mer (soit près d’un an), pour essayer de mieux comprendre la distribution et les trajectoires de ces oiseaux marins.
Pour pouvoir retranscrire les mouvements en mer des individus, les chercheurs fixent des émetteurs reliés au système de géolocalisation Argos [**] sur le dos des oiseaux (plus d’informations dans cette vidéo du CNES). L’émetteur contient aussi un capteur de pression qui permet de mesurer la profondeur à laquelle se trouve le manchot et de reconstruire le profil de plongée. Afin de collecter des données sur plusieurs mois, les chercheurs ont dû faire un compromis entre la durée de vie de la batterie et la quantité de données récoltées. Ainsi, les « tags » (émetteur + capteur) enregistrent les données de localisation (latitude et longitude) toutes les deux heures, et la pression un jour sur trois (soit toutes les secondes pendant 24 h pour une période de 72 h).
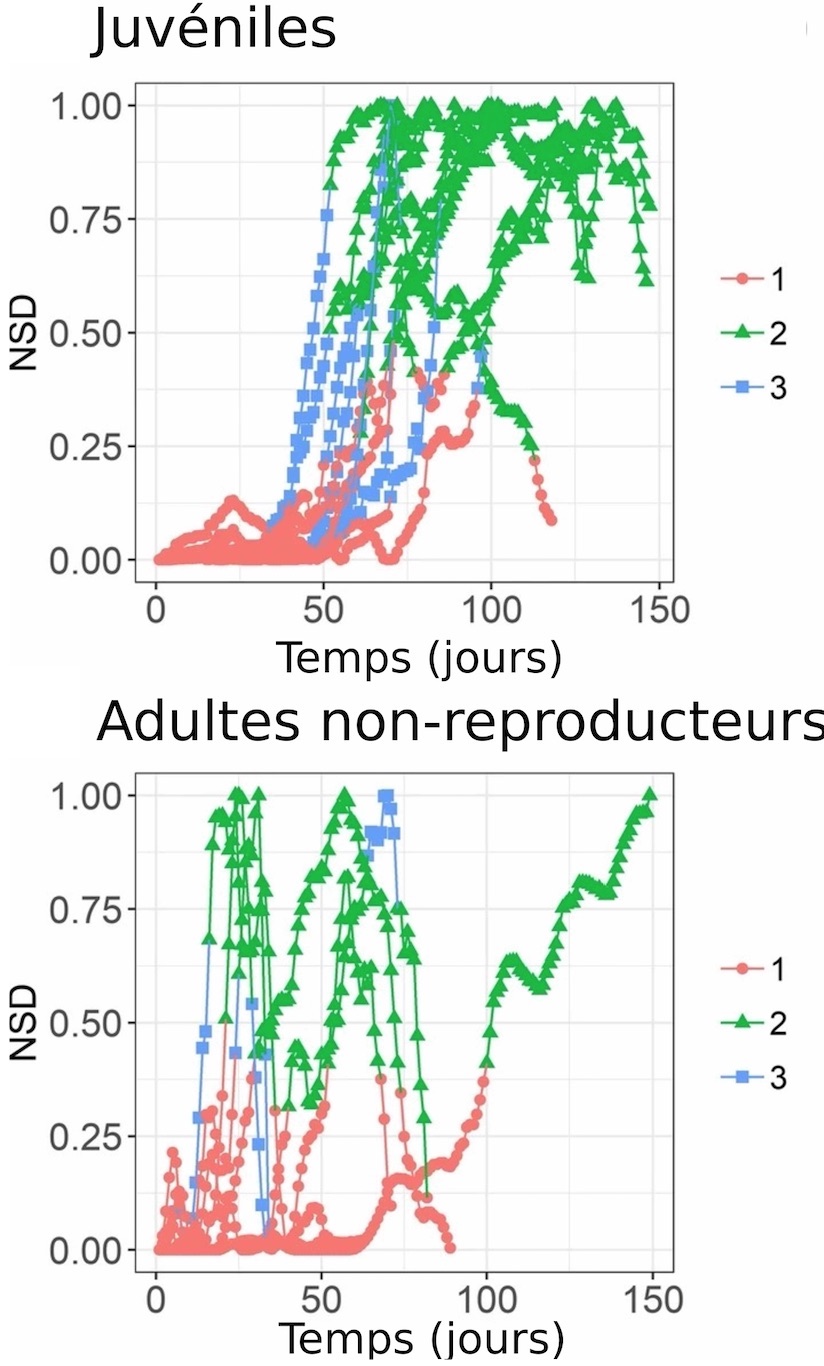
À partir des données de localisations Argos, il est possible d’obtenir le NSD (Net Squared Displacement, soit « déplacement net au carré ») en calculant la distance entre chacune des positions d’un manchot et son lieu de départ. Comme chaque position correspond à un temps donné, on peut retracer un profil NSD en fonction du temps passé en mer (Figure 1). Cette analyse permet ensuite de caractériser les différentes phases du mouvement et notamment de distinguer les zones géographiques que les manchots vont simplement traverser de celles dans lesquelles ils passent plus de temps pour explorer ou se nourrir. En utilisant cette technique, les chercheurs ont pu séparer le mouvement en 3 phases : les phases 1 et 2 correspondent à des mouvements regroupés dans des zones géographiques précises (probablement des sites où les manchots se nourrissent), respectivement en été et en hiver (Figure 1, ronds rouges et triangles vert), et la troisième phase correspond à la transition entre ces deux aires géographiques (Figure 1, carrés bleus). Les chercheurs ont également extrait des variables environnementales relatives aux lieux fréquentés par les manchots, afin de mettre en lumière les conditions environnementales recherchées pour trouver de la nourriture.
Les détails du voyage : où et quand partent les manchots ?
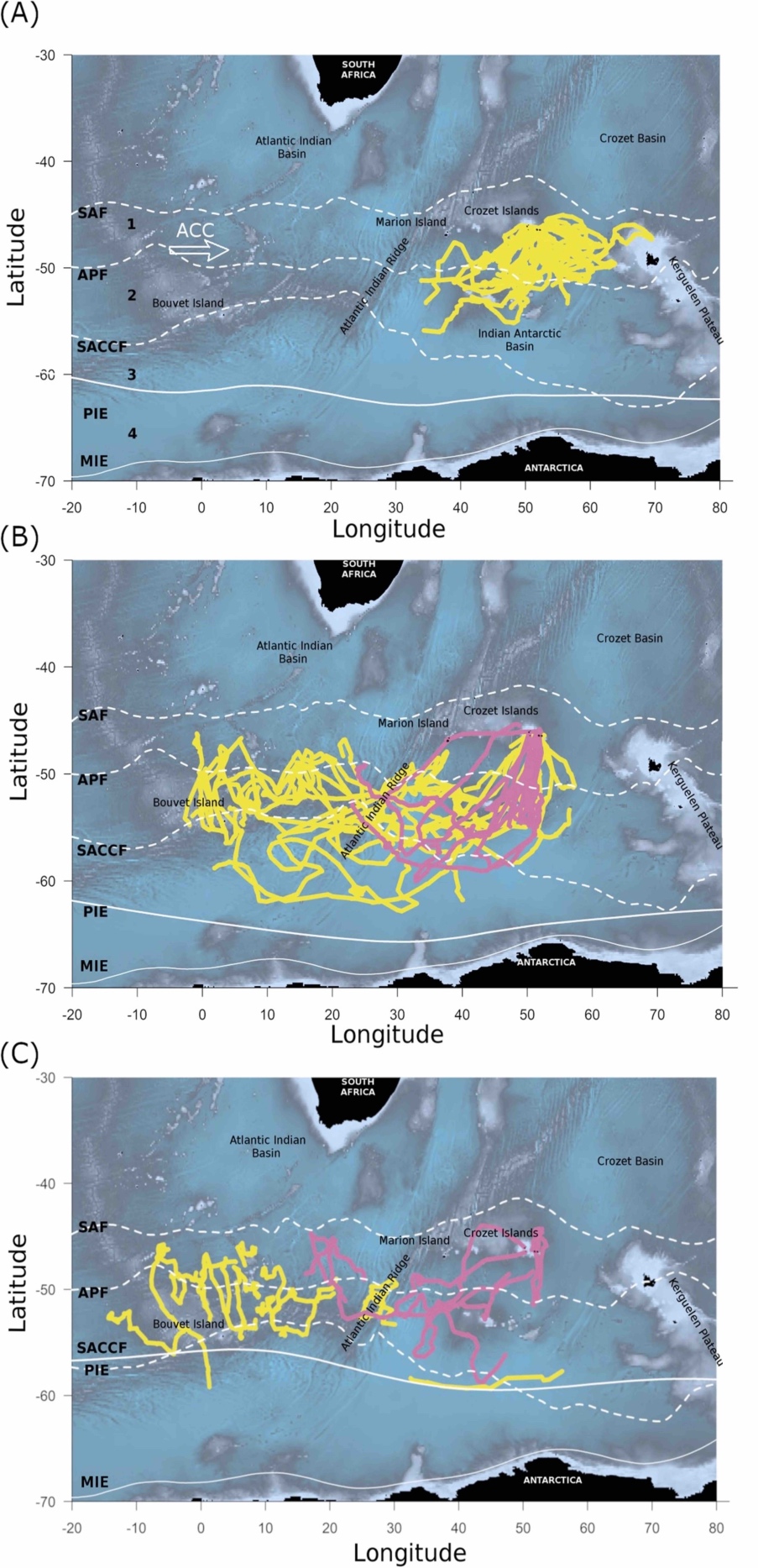
Été austral (Figure 2, A)
Durant l’été, les juvéniles explorent de larges aires géographiques au sud de la colonie, mais sont principalement retrouvés dans la Zone Frontale Polaire, à proximité du Front Antarctique Polaire (APF ; voir Figure 2). Cette zone est connue comme étant un réservoir important de nourriture pour les manchots de cette région. Il est donc à envisager que les juvéniles suivent les adultes reproducteurs qui se nourrissent aux alentours de la colonie pendant l’été.
Les adultes non-reproducteurs ne quittent l’île qu’à la mi-février, voire en mars, après avoir terminé leur mue annuelle.
Automne (Figure 2, B)
Après une exploration estivale au sud de l’archipel de Crozet, les juvéniles commencent à se diriger plus loin vers le sud-ouest au mois de mars. Ce déplacement peut être lié à une diminution de l’abondance des ressources dans la Zone Frontale Polaire. Les juvéniles partent à la recherche de zones d’alimentation où la compétition intra-spécifique — c’est-à-dire avec des congénères de leur propre espèce — pour les ressources est moins forte, les aires d’alimentation aux alentours de l’île étant également utilisées par les adultes reproducteurs. Les adultes non-reproducteurs plus expérimentés suivent la même stratégie plus tard dans la saison.
Hiver (Figure 2, C)
Les manchots atteignent une zone géographique lointaine au sud-ouest par rapport à l’île de la Possession, mais se répartissent sur une aire assez vaste. Les jeunes manchots rejoignent cette zone plus tôt que les adultes non-reproducteurs, ce qui suggère qu’ils sont capables de trouver les zones intéressantes pour passer l’hiver sans apprentissage préalable avec des congénères. L’analyse des dates de départ et d’arrivée des manchots révèle une grande variabilité entre les individus (juvéniles ou adultes). De la même façon, l’analyse des paramètres environnementaux (température des eaux de surface, présence et concentration de glace de mer, vitesse des vents et courants, etc.) montre que les manchots traversent une grande diversité d’environnements pendant l’hiver. Ces résultats ne vont pas dans le sens d’un mouvement migratoire stricto sensu pour lequel on s’attendrait plutôt à un départ simultané des individus vers des zones géographiques bien définies [8]. Les auteurs en déduisent que les mouvements saisonniers des manchots relèvent plutôt d’un mélange entre des mouvements migratoires et exploratoires.
Une détermination innée du mouvement…
Les données de positions Argos des manchots indiquent que, sur la totalité de l’étude, les jeunes manchots et les adultes non-reproducteurs se répartissent sur les mêmes latitudes avec une limite nord identique. En revanche, les jeunes manchots ont atteint des limites sud et ouest plus lointaines que celles atteintes par les autres adultes. Ce résultat indique que les juvéniles ont un comportement plus exploratoire que les adultes expérimentés, qui semblent cibler plus précisément certaines zones. Les adultes plus âgés comptent souvent se reproduire l’année suivant une année sans reproduction, donc il est possible qu’ils s’éloignent moins de leur colonie d’origine afin de pouvoir y retourner rapidement à la bonne saison. Le fait que les juvéniles couvrent des distances plus grandes, et qu’ils arrivent plus tôt que les adultes non-reproducteurs, indique qu’ils sont capables de rejoindre des zones d’alimentation importantes par eux-mêmes. Ainsi, les mouvements des juvéniles sont synchrones et dirigés (tous les individus vont plus ou moins dans la même direction et les changements de direction ou de types de mouvement ont lieu en même temps) ce qui suggère que le déplacement saisonnier des manchots serait (au moins en partie) le résultat d’un comportement inné de dispersion propre aux manchots.
Cependant, dans un environnement en constante évolution et souvent imprédictible, il est difficile de croire que seule une programmation génétique de la navigation puisse suffire, même en prenant en compte la variabilité dans l’information génétique et sa transmission. Être capable d’adopter des comportements différents suivant les conditions environnementales dans lesquelles ils se trouvent pourrait permettre aux manchots de faire face à un environnement changeant (déplacement des zones avec beaucoup de proies, présence de glace, modification des courants, etc.). On dit que leur comportement est « plastique », c’est-à-dire qu’il change en fonction des conditions du milieu rencontrées. On peut donc suspecter qu’il existe d’autres mécanismes permettant aux manchots d’optimiser leurs déplacements vers des zones riches en ressources, et ainsi de se nourrir plus efficacement pour conserver une balance énergétique positive, cruciale pour leur survie.
… et l’influence de l’environnement
Afin de mieux comprendre ce qui détermine les mouvements des manchots royaux en mer, les chercheurs ont voulu comparer l’orientation des individus par rapport à plusieurs paramètres environnementaux comme la direction des vents, des courants ou la productivité océanique des zones traversées (représentée ici par la concentration en chlorophylle a, notée « chl a »). Ces données environnementales proviennent de différentes bases de données obtenues grâce aux données collectées par des satellites [***]. L’orientation des manchots est calculée comme la différence d’angle entre la direction du courant ou du vent et la direction de déplacement du manchot (obtenue à partir des positions Argos). Pour simplifier les données, les chercheurs ont réparti les valeurs en 3 catégories : même direction (valeurs entre 315° et 45°), direction opposée (entre 135° et 225°), et directions perpendiculaires pour toutes les autres valeurs (Figure 3). Ainsi, pour chaque position relevée pour un individu, l’orientation par rapport au vent et au courant est connue.
Les chercheurs se sont demandé si l’orientation des manchots par rapport au vent et au courant pouvait dépendre de la productivité océanique (c’est à dire la productivité primaire dans les océans qui est liée à la photosynthèse réalisée par le phytoplancton, il s’agit de la base de la chaîne alimentaire dans les océans). Pour cela, ils ont comparé l’orientation des manchots à l’instant t par rapport au vent et au courant suivant les valeurs de productivité océanique à leurs positions précédentes (t-1) et suivante (t+1 ; Figure 4). Cela permet ainsi d’estimer si l’orientation des manchots est préférentiellement dirigée selon un gradient de productivité océanique ou selon la direction « par défaut » imposée par le vent ou les courants marins. Autrement dit, les manchots sont-ils capables de détecter les zones intéressantes en termes de ressources alimentaires et de s’y rendre activement ?
Sur la totalité de l’étude, les manchots se sont déplacés le plus souvent perpendiculairement au vent et au courant (environ 50 % des mesures correspondent à la catégorie perpendiculaire). Quand la direction des manchots n’est pas perpendiculaire au vent ou au courant, ces derniers ont adopté à la fois des directions opposées ou dans le même sens que le vent et le courant selon les saisons : ils nagent le plus souvent dans le sens contraire aux vents et aux courants en automne, et dans le même sens en hiver. De plus, les analyses statistiques ont montré que les manchots ont davantage tendance à s’orienter contre le sens du vent si la concentration en chlorophylle a est supérieure à leur prochaine position par rapport à leur précédente (Figure 5A). Cet effet n’est pas observé dans le cas du courant.
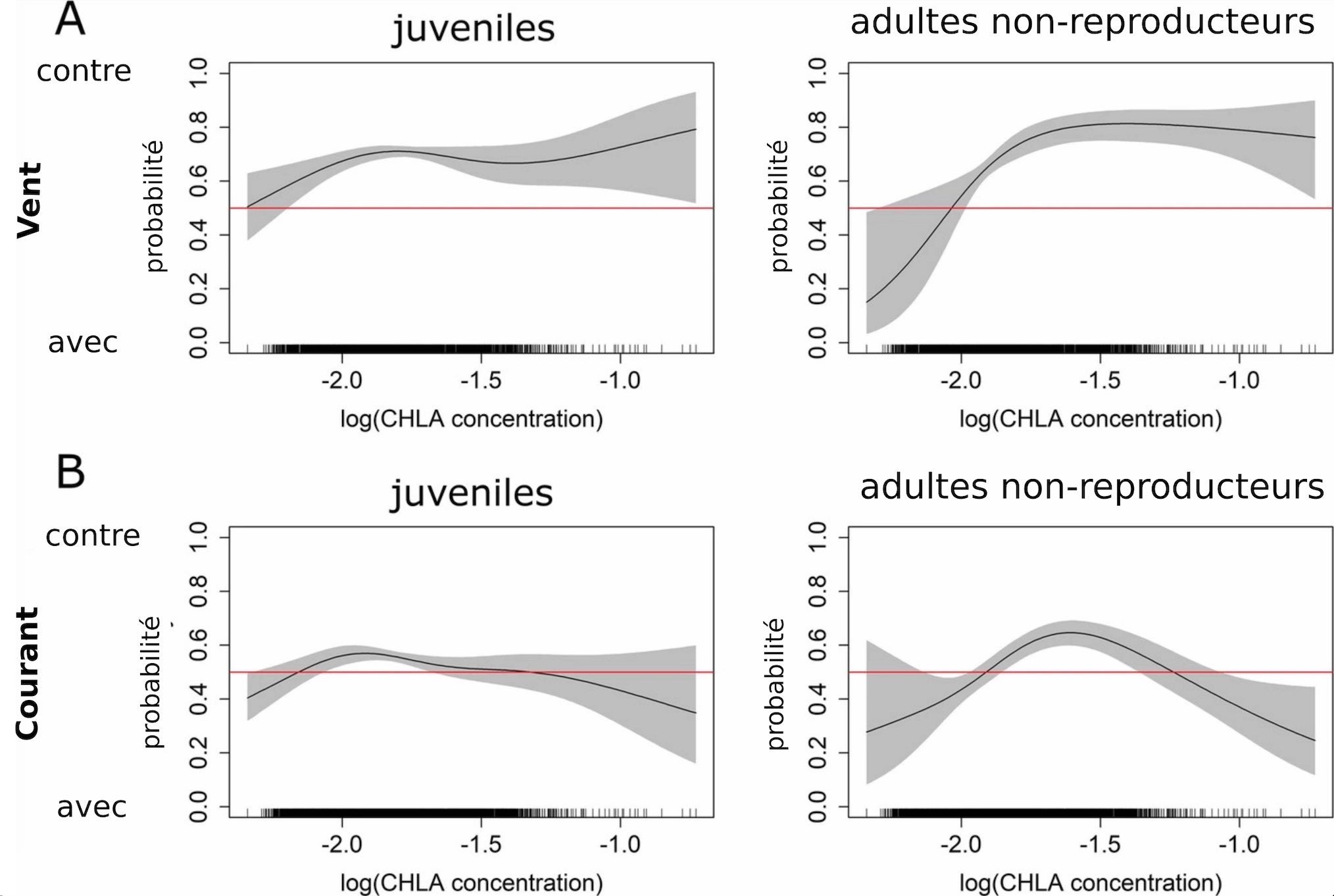
Ces résultats suggèrent que les manchots montrent des orientations préférentielles par rapport aux vents et aux courants et que ces orientations dépendent d’autres paramètres comme la saison ou la productivité océanique. Il semblerait notamment que le vent soit un paramètre crucial. En revanche, les auteurs doutent que les manchots soient capables de ressentir le courant, et donc de s’orienter par rapport à ce paramètre. En effet, ces oiseaux marins n’ont pas de repère stationnaire (fixe) aux profondeurs auxquelles ils plongent (comme pourrait l’être le plancher océanique). Ainsi, les auteurs concluent que les orientations observées par rapport au courant sont probablement dues à une coïncidence. On peut aussi penser que les résultats sur le courant découlent de l’orientation des manchots par rapport au vent puisque les directions du vent et du courant sont intimement corrélées (au moins en surface) [9]. Le résultat le plus frappant concerne l’orientation des manchots par rapport au vent quand on considère la productivité océanique en amont et en aval de leur position. Ce résultat suggère que les manchots sont capables de s’orienter vers des zones riches dans le sens opposé au vent. Cela impliquerait que le vent transporte des molécules en provenance de ces zones, que celles-ci seraient détectables par les manchots et que ces derniers remonteraient la piste jusqu’à ces aires productives. Cette hypothèse est plausible puisque des phénomènes similaires ont été observés chez d’autres espèces (comme les tortues [10]). Les manchots reviennent souvent à la surface pour respirer et pourraient donc inhaler ces molécules. Cependant, un manque d’information subsiste quant aux capacités olfactives des manchots et de nouvelles études seraient nécessaires afin d’estimer dans quelles mesures ils pourraient utiliser ces indices pour s’orienter.
Conclusion
Dans cette étude, les chercheurs ont pu mettre en lumière les comportements de déplacement des manchots royaux lors de deux phases peu connues de leur cycle de vie : lors du premier départ en mer des juvéniles et lors d’un voyage en mer des adultes non-reproducteurs. Grâce aux techniques de télémétrie et d’analyse des données collectées, les chercheurs ont pu analyser le mouvement des manchots à plusieurs échelles spatiales et temporelles. Sur toute la durée de l’étude, et à une large échelle spatiale, il ressort que le mouvement des manchots peut se découper en 3 phases, qu’ils ne rejoignent pas d’aire géographique particulière et que les juvéniles ont un comportement plus exploratoire que les adultes non-reproducteurs. Ces résultats suggèrent que les manchots royaux ne sont pas des oiseaux migrateurs à proprement parler, même s’il semble y avoir une composante innée dans les choix qui déterminent leurs mouvements saisonniers.
En regardant à une échelle plus locale, les manchots semblent utiliser des indices environnementaux présents dans leur milieu, et notamment la direction du vent, pour naviguer et trouver des zones riches en ressources alimentaires. Cette utilisation d’informations environnementales leur permettrait d’être plus efficaces dans leur réapprovisionnement et en meilleure condition corporelle. Cette compétence est cruciale car le réapprovisionnement en mer est une étape clé dans le cycle de vie du manchot et détermine pour beaucoup à la fois la survie des individus et leurs chances de réussite pour la prochaine reproduction.
Pour aller plus loin, il serait intéressant de poursuivre l’étude en s’intéressant à d’autres phases du cycle de vie des manchots, notamment en comparant les mouvements des individus entre leur premier voyage en mer et les suivants. Cela permettrait de mieux saisir les facteurs qui influencent la transition du comportement de navigation depuis la phase juvénile jusqu’à l’âge adulte et notamment de mieux comprendre l’importance relative qu’occupe l’apprentissage de ces comportements par rapport au renforcement de comportements innés. La poursuite de ces travaux permettrait de façon plus générale de mieux comprendre comment des comportements adaptés à des environnements et des contraintes spécifiques à cette espèce ont pu voir le jour et perdurer dans les populations de manchots.
[*] Photographie également publiée dans l’ouvrage Aventures Australes aux éditions Omniscience.
[**] Le système ARGOS (Advanced Research and Global Observation Satellite) est un système satellitaire de localisation comparable au système GPS (Global Positioning System), mais il fonctionne différemment et ne donne pas les positions avec la même précision. Les balises ARGOS transmettent les données de localisation à l’aide d’un effet Doppler, alors que les balises GPS collectent et stockent les données de localisation. Pour plus d’informations sur les balises ARGOS et leur intérêt pour l’étude des manchots, vous pouvez consulter cette vidéo du CNES. Vous pouvez également lire ce papier mâché sur une autre utilisation possible du système ARGOS.
[***] Les données proviennent du Global Forecast System of the United States National Weather Service et du Copernicus Marine Environment Monitoring Service of the European Union.
[1] Lohmann K. J., et al., The sensory ecology of ocean navigation. Journal of Experimental Biology, 2008. DOI : 10.1242/jeb.015792. [Publication scientifique]
[2] Bost C. A., et al., Large-scale climatic anomalies affect marine predator foraging behaviour and demography. Nature Communications, 2015. DOI : 10.1038/ncomms9220. [Publication scientifique]
[3] Cherel Y. & Ridoux V.. Prey species and nutritive value of food fed during summer to King Penguin Aptenodytes patagonica chicks at Possession Island, Crozet Archipelago. Ibis, 1992. DOI : 10.1111/j.1474-919X.1992.tb08388.x. [Publication scientifique]
[4] Charrassin J.-B. & Bost C. A., Utilisation of the oceanic habitat by king penguins over the annual cycle. Marine Ecology Progress Series, 2001. DOI : 10.3354/meps221285. [Publication scientifique]
[5] Froget G., et al., Heart rate and energetics of free-ranging king penguins (Aptenodytes patagonicus). Journal of Experimental Biology, 2004. DOI : 10.1242/jeb.01232. [Publication scientifique]
[6] Enstipp M. R., Apparent changes in body insulation of juvenile king penguins suggest an energetic challenge during their early life at sea. Journal of Experimental Biology, 2017. DOI : 10.1242/jeb.160143. [Publication scientifique]
[7] Clucas G. V., et al., Dispersal in the sub-Antarctic: king penguins show remarkably little population genetic differentiation across their range. BMC Evolutionary Biology, 2016. DOI : 10.1186/s12862-016-0784-z. [Publication scientifique]
[8] Bauer S., et al., Timing is crucial for consequences of migratory connectivity. Oikos, 2016. DOI : 10.1111/oik.02706. [Publication scientifique]
[9] Song H., et al., Impact of Current-Wind Interaction on Vertical Processes in the Southern Ocean. Journal of Geophysical Research: Oceans, 2020. DOI : 10.1029/2020JC016046. [Publication scientifique]
[10] Galli S., et al., Orientation of migrating leatherback turtles in relation to ocean currents. Animal Behaviour, 2012. DOI : 10.1016/j.anbehav.2012.09.022. [Publication scientifique]
Publié le 19/10/2021
Camille Lemonnier/Papier-Mâché/CC BY-NC-SA 4.0 2021Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.