Chargement de l'article...
Écriture : Élodie Billard
Relecture scientifique : Anna-Margarete Stahler
Relecture de forme : Jérémy Ferrand et Eléonore Pérès
Temps de lecture : environ 10 minutes.
Thématiques : Évolution et Microbiologie (Biologie)
Publication originale : Imachi H., et al., Isolation of an archaeon at the prokaryote–eukaryote interface. Nature, 2020. DOI : 10.1038/s41586-019-1916-6
Vous vous êtes peut-être déjà demandé où et quand était apparue l’espèce humaine. Qui de l’œuf ou de la poule était venu en premier. Mais vous êtes-vous déjà demandé d’où viennent les cellules qui nous composent ? Comment sont-elles apparues sur Terre ? La question de l’apparition des Eucaryotes n’est toujours pas résolue et fait débat parmi les scientifiques. Toutefois, une récente étude met en lumière un microorganisme et des processus encore mal compris jusqu’alors qui pourraient bien nous permettre d’y voir plus clair et de répondre enfin à cette question.
Il était une fois, il y a très très longtemps, une planète hostile où l’oxygène était rare et qui abritait des organismes unicellulaires simples comme seuls habitants. Bienvenue sur Terre il y a environ 3 milliards d’années…
La vie commence à s’installer progressivement sur Terre il y a environ 3,8 milliards d’années, suite à l’apparition de l’oxygène, gaz essentiel à la vie de la plupart des organismes actuels. La Terre est alors constituée d’un océan recouvrant quasiment toute la planète et d’une atmosphère primitive, différente de ce qu’elle est aujourd’hui [1]. Les organismes vivants sont alors adaptés à une vie marine : un milieu pauvre en oxygène et un rayonnement ultraviolet important provenant du Soleil, et non filtré par l’atmosphère de l’époque. Sous l’effet de la multiplication des microorganismes — qui rejettent l’oxygène comme un déchet — et de modifications de la composition des roches de l’enveloppe terrestre, l’oxygène va s’accumuler dans l’atmosphère [2]. Puis, il y a environ 2,3 milliards d’années, la quantité d’oxygène augmente fortement en peu de temps (environ 10 millions d’années, soit l’équivalent de 8 secondes dans 1 heure, vous voyez que c‘est court !). Cet événement est appelé grande oxygénation (ou oxydation) de l’atmosphère terrestre. Pour les organismes vivants, adaptés à une vie pauvre en oxygène depuis presque 1 milliard d’années, la forte augmentation de la concentration en oxygène va avoir des effets toxiques. Ils vont devoir s’adapter à ce changement important. C’est dans ce contexte qu’apparaissent, il y a environ 2 milliards d’années, des formes de vie diverses et plus complexes : les Eucaryotes.
Kesako les Eucaryotes ?
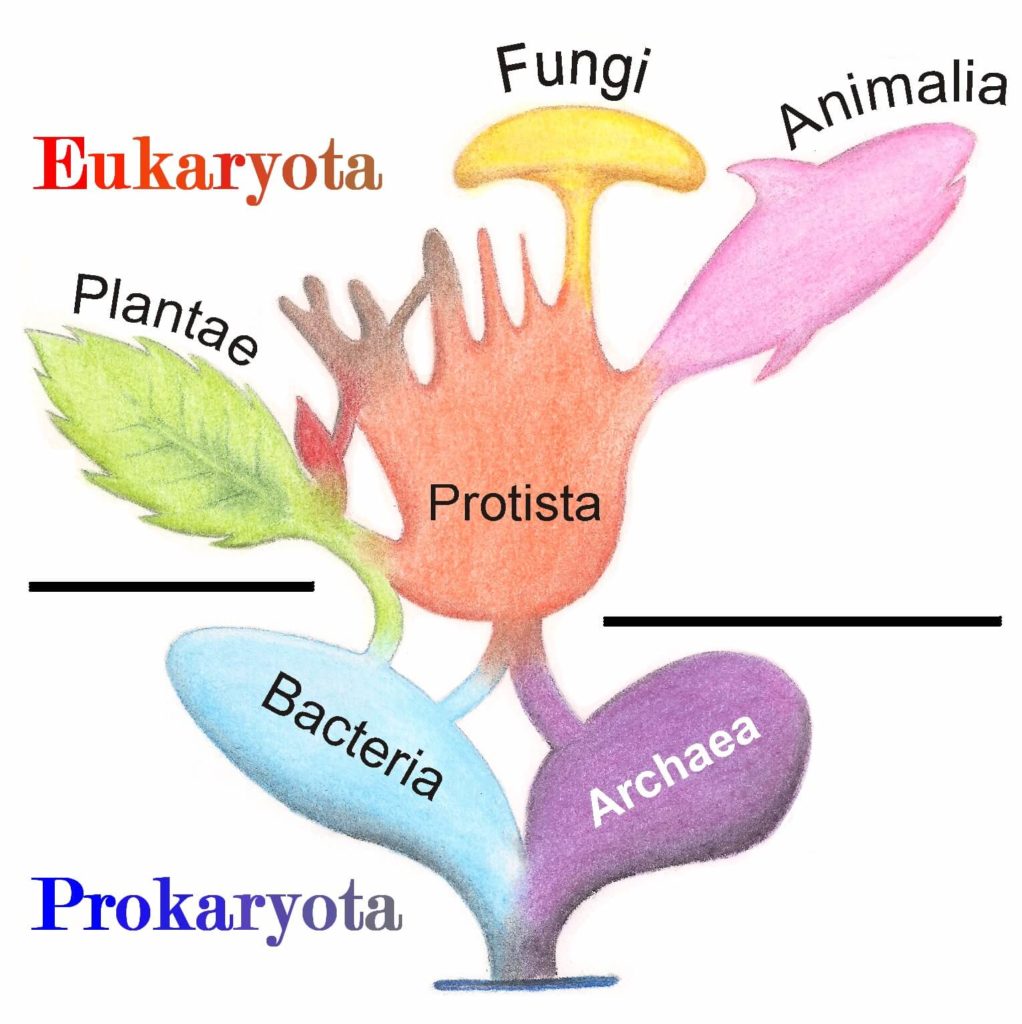
L’ensemble des organismes vivants sur Terre est actuellement classé en trois domaines : les Archées, les Bactéries et les Eucaryotes (Figure 1). Le domaine des Archées est le moins connu des trois, et pour cause, pendant longtemps les Archées ont été assimilées au domaine des Bactéries, alors appelées Archéobactéries. C’est en 1990 que Carl Woese et ses collaborateurs [3] vont insister sur le fait que les « archéobactéries » sont un groupe fondamentalement différent des bactéries. Il crée alors le domaine des Archées. Celui des Eucaryotes comprend l’ensemble des animaux (Animalia, dont nous faisons partie) ainsi que les plantes (Plantae) et les champignons (Fungi) (Figure 1).
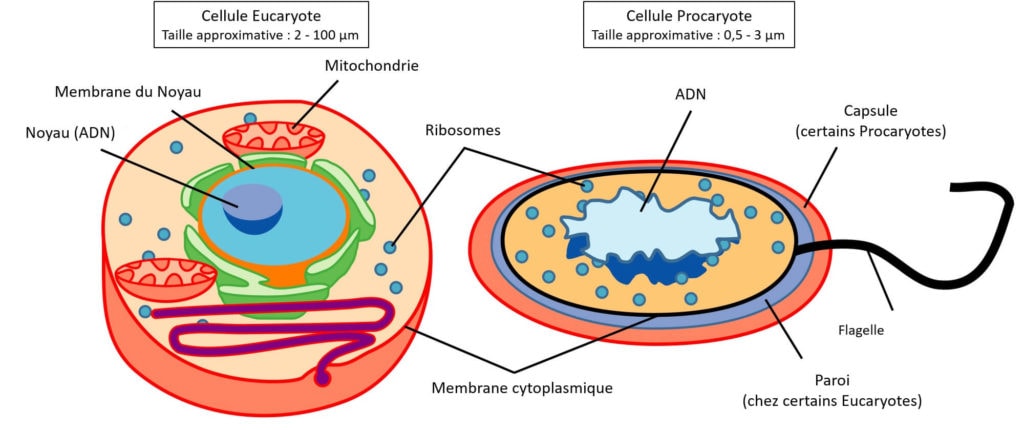
Les cellules eucaryotes sont considérées comme étant plus complexes que celles des Bactéries et des Archées. Elles présentent des structures caractéristiques qui ne sont pas partagées avec les Procaryotes (Bactéries et Archées) (Figure 2). Le nom Eucaryote signifie « ceux qui possèdent un noyau » (du grec ancien ευ- (« eu ») signifiant « bien » et -καρυο𝜈 (« karuon ») pour « noyau »), s’opposant ainsi aux Procaryotes qui n’en possèdent pas. La présence d’autres structures spécialisées délimitées par une membrane (par exemple : la mitochondrie comme présenté Figure 2) est aussi une caractéristique des Eucaryotes.
Une question qui taraude les scientifiques depuis longtemps est de comprendre comment ces cellules « complexes » sont apparues il y a environ 2 milliards d’années, soit environ 1 milliard d’années après l’apparition de la vie sur Terre.
L’hypothèse la plus couramment acceptée actuellement est celle de la théorie endosymbiotique. Selon cette théorie, certaines structures spécialisées caractéristiques des cellules eucaryotes (comme les mitochondries, voir Figure 2) proviendraient de l’incorporation d’une cellule procaryote par une autre. On pourrait dire qu’une cellule a « ingurgité » puis « hébergé » une autre cellule et a ensuite converti cette dernière en structure interne pour son propre compte. Pour que cette théorie puisse se tenir, il nous faut deux acteurs :
- une cellule hôte (qui héberge un autre organisme) : probablement une archée qui serait habituée à la vie sans ou avec très peu d’oxygène ;
- une cellule hébergée (dite endosymbiote) : probablement une bactérie, tolérante à l’oxygène.
Une relation de bénéfices mutuels se serait alors mise en place entre les deux organismes à la suite de cette « incorporation », appelée endosymbiose : l’organisme « hébergé » procurant de l’énergie et une tolérance à l’oxygène à l’hôte, en échange d’un habitat protégé et d’un échange mutuel de nutriments.
Nous avons des preuves scientifiques de cette théorie, comme la présence d’un ADN propre à certaines structures cellulaires : les mitochondries par exemple possèdent un ADN particulier, différent de l’ADN de la cellule eucaryote. Pour autant, nous ne savons toujours pas précisément qui sont les deux acteurs à l’origine des premières cellules eucaryotes. Ceci est toujours un sujet de débats au sein de la communauté scientifique. Depuis plusieurs années, l’identité de l’endosymbiote a commencé à s’éclaircir : il s’agirait d’une espèce de bactérie appartenant aux Alpha-Protéobactéries. Mais jusqu’alors, l’hôte restait inconnu chez les Archées…
Nom de code : MK-D1
La publication d’Imachi et de ses collaborateurs concernant la découverte d’une nouvelle espèce d’archées par une équipe de chercheurs japonais vient d’apporter un nouvel éclairage sur les évènements ayant conduit à l’apparition des Eucaryotes sur Terre. En effet, cette nouvelle archée pourrait être justement l’hôte impliqué dans la théorie endosymbiotique !
Les archées qui nous intéressent ici forment un groupe particulier, du nom de groupe d’Asgård. La plupart des noms des archées de ce groupe est basé sur des dieux de la mythologie nordique. Ainsi on trouve : Odin-, Thor-, Loki-, Hel- et Heimdallarchaeota.
C’est au sein des Lokiarchaeota — nous les appellerons Loki pour simplifier — que se trouve l’archée qui nous intéresse dans cette publication. L’histoire commence en 2006, lorsqu’une équipe de chercheurs japonais réalise des prélèvements de sédiments marins sur la crête d’Omine, dans la fosse de Nankai au Japon, à plus de 2 500 mètres de profondeur. Les premières analyses révèlent assez vite la présence de nombreux microorganismes encore inconnus. L’équipe décide alors d’essayer de cultiver ces microorganismes inconnus en laboratoire. Mais ils font alors face à plusieurs défis :
- les archées, en général, sont difficilement cultivables en laboratoire ;
- l’échantillon collecté provient d’une zone sans oxygène et riche en méthane : il faut donc reproduire ces conditions.
Après plus de 2 000 jours (soit presque 5 ans et demi !) de culture en laboratoire, l’équipe réussit à détecter la présence de microorganismes divers. Afin de les isoler pour mieux les étudier, l’équipe poursuit les expériences. C’est ainsi qu’après plus de 10 années de recherche, l’équipe identifie finalement une archée encore inconnue, appelée dans un premier temps MK-D1 (Figure 3).

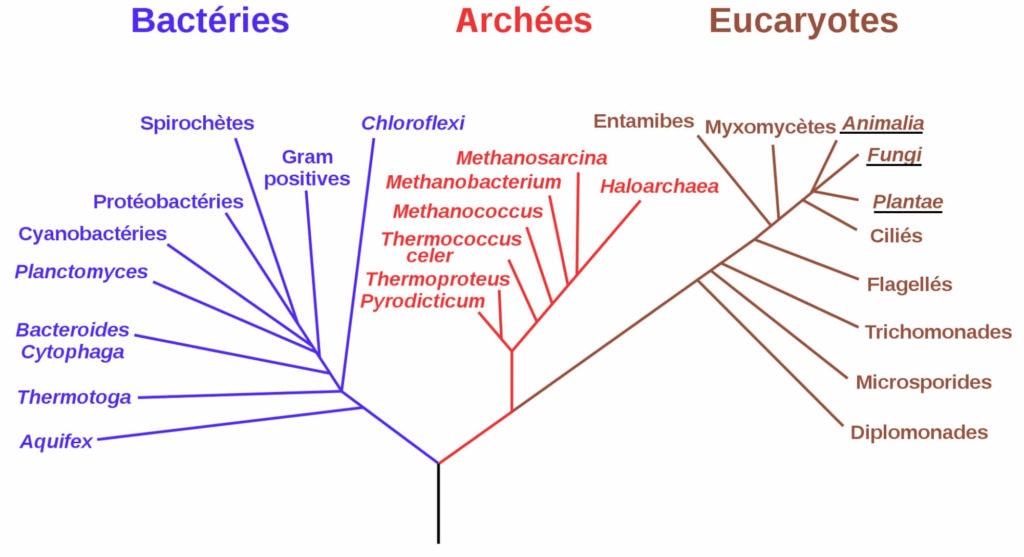
La découverte de cette nouvelle archée est déjà un événement en soi. Mais certaines caractéristiques de MK-D1 ont alors suscité la curiosité des scientifiques de l’étude et les ont conduits à pousser leur réflexion plus loin. En effet, cette archée vit dans des milieux où l’oxygène est rare voire absent, des milieux semblables aux conditions de vie d’il y a environ 2 milliards d’années. Elle produit des « bras » ou « tentacules » qui lui permettent de se lier à d’autres microorganismes et donc éventuellement d’en « avaler » un autre. Mais le plus intéressant, c’est que cette archée est l’individu le plus proche génétiquement des Eucaryotes (Figure 4). Ce nouvel organisme réunit donc certaines conditions qui permettent d’émettre l’hypothèse qu’il pourrait être un hôte possible, jusqu’à alors encore inconnu, de la théorie endosymbiotique à l’origine de l’émergence des Eucaryotes sur Terre.
Mission : Enchevêtrer – Engloutir – Internaliser
À partir des caractéristiques identifiées chez la nouvelle archée MK-D1 et en partant de l’hypothèse que celle-ci pourrait être un hôte possible pour la théorie endosymbiotique, les auteurs de la publication ont établi un nouveau modèle nommé E3 pour Enchevêtrement – Engloutissement – Endogénéisation (= internalisation).
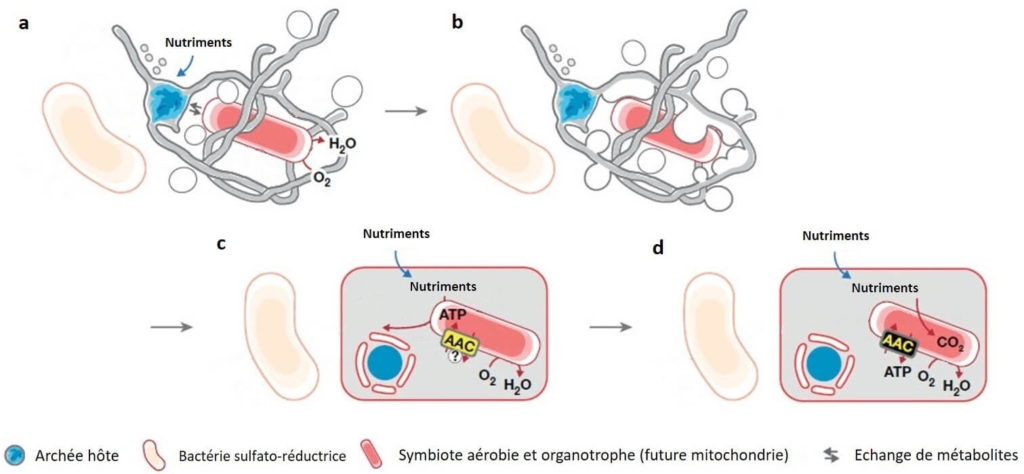
Ils ont alors proposé un récit des événements qui auraient amené MK-D1 à « avaler » un autre organisme et pouvant expliquer l’origine des Eucaryotes. Selon les auteurs, plusieurs étapes auraient été nécessaires (Figure 5).
- La première étape est une transition entre un mode de vie sans oxygène vers un mode de vie tolérant la présence d’oxygène. Une relation de dépendance se serait installée entre les deux organismes : l’archée, futur hôte et la bactérie, futur endosymbiote (Figure 5a).
- Ensuite, les structures externes de l’hôte (« bras ou tentacules », Figure 3) auraient interagi avec la bactérie, permettant d’améliorer l’interaction physique et « d’engloutir » celle-ci. On pourrait dire que l’hôte a « avalé » la bactérie mais sans la « digérer ». (Figure 5b).
- Suite à cet « engloutissement », l’hôte aurait partagé avec la bactérie symbiotique des molécules comme source d’énergie pour cette dernière. En retour, la bactérie aurait consommé l’oxygène environnant et fourni à son tour de l’énergie à l’hôte sous forme de la molécule ATP (Figure 5c).
- La symbiose entre les deux organismes aurait évolué au cours du temps. Ainsi, à terme, l’hôte aurait délégué la production d’énergie sous forme d’ATP à la bactérie. Un canal, appelé AAC, qui permet le passage de l’ATP entre les deux cellules se serait mis en place (Figure 5d). Ce canal est aujourd’hui essentiel pour les mitochondries actuelles des cellules eucaryotes.
- Finalement, au cours de l’évolution, des modifications dans la structure interne seraient survenues, donnant naissance aux mitochondries actuelles. Il est aussi possible que d’autres endosymbioses se soient produites en série pour parvenir aux Eucaryotes tels que nous les connaissons aujourd’hui.
Ce modèle et l’enchaînement des évènements décrit ici n’est qu’un scénario hypothétique parmi d’autres pour expliquer l’émergence des Eucaryotes sur Terre. Il est basé sur les connaissances scientifiques actuelles. C’est tout l’intérêt de cette publication, les travaux décrits ici permettent d’améliorer nos connaissances. D’une part, par la découverte d’un nouvel organisme et la caractérisation de son métabolisme et de sa physiologie et d’autre part, par le développement d’une nouvelle théorie pouvant répondre à une question qui fait débat depuis longtemps au sein de la communauté scientifique. C’est la poursuite des recherches dans les années à venir qui permettra d’évaluer la validité du scénario décrit ici. Mais aussi, peut-être, de répondre à des questions encore en suspens à ce jour sur l’engloutissement de la bactérie et son internalisation et enfin sur l’enchaînement des évènements eux-mêmes.
[1] Johnson B.W. & Wing B.A., Limited Archaean continental emergence reflected in an early Archaean 18O-enriched ocean. Nature Geoscience, 2020. DOI : 10.1038/s41561-020-0538-9. [Publication scientifique]
[2] Smit M. & Mezger K., Earth’s early O2 cycle suppressed by primitive continents. Nature Geoscience, 2017. DOI : 10.1038/ngeo3030. [Publication scientifique]
[3] Woese C.R., et al.,Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. PNAS, 1990. DOI : 10.1073/pnas.87.12.4576. [Publication scientifique]
Publié le 21/12/2020
 Elodie Billard/Papier-Mâché/CC BY-NC-SA 4.0 2020
Elodie Billard/Papier-Mâché/CC BY-NC-SA 4.0 2020Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.
Écriture : Élodie Billard
Relecture scientifique : Anna-Margarete Stahler
Relecture de forme : Jérémy Ferrand et Eléonore Pérès
Temps de lecture : environ 12 minutes.
Thématiques : Évolution et Microbiologie (Biologie)
Publication originale : Imachi H., et al., Isolation of an archaeon at the prokaryote–eukaryote interface. Nature, 2020. DOI : 10.1038/s41586-019-1916-6
Vous vous êtes peut-être déjà demandé où et quand était apparue l’espèce humaine. Qui de l’œuf ou de la poule était venu en premier. Mais vous êtes-vous déjà demandé d’où viennent les cellules qui nous composent ? Comment sont-elles apparues sur Terre ? La question de l’apparition des Eucaryotes n’est toujours pas résolue et fait débat parmi les scientifiques. Toutefois, une récente étude met en lumière un microorganisme et des processus encore mal compris jusqu’alors qui pourraient bien nous permettre d’y voir plus clair et de répondre enfin à cette question.
Comment sont apparus les Eucaryotes ?
Les Eucaryotes sont l’un des trois domaines du vivant avec les Bactéries et les Archées (Figure 1). Comme leur nom l’indique, les Eucaryotes sont des organismes caractérisés par la présence d’un noyau dans leurs cellules : ευ- (« eu ») du grec « bien » et -καρυο𝜈 (« karuon ») pour « noyau ». Cela signifie « ceux qui possèdent un noyau », s’opposant ainsi aux Procaryotes qui n’en ont pas. Les Procaryotes regroupent les deux autres domaines du vivant : les Bactéries et les Archées mais ne forment pas un taxon admis du point de vue phylogénétique. De plus, les Eucaryotes sont pourvus d’organites spécialisés tels que les mitochondries et les chloroplastes, ils ont donc des caractéristiques cellulaires plus complexes que les Procaryotes. Parmi les Eucaryotes, on retrouve les animaux (Animalia, dont nous faisons partie), les végétaux et les champignons (Fungi) par exemple (Figure 1).
Pour bien comprendre dans quel contexte sont apparus les Eucaryotes, il faut se remémorer les conditions environnementales sur Terre à ce moment-là. Il y a environ 3 milliards d’années, l’oxygène a commencé à s’accumuler sur Terre, d’abord dans les océans puis dans l’atmosphère, principalement grâce à la multiplication de microorganismes photosynthétiques et de phénomènes géophysiques [1]. L’oxygène est alors rejeté par les microorganismes, il s’agit d’un déchet de leurs réactions métaboliques. Puis vers 2,3 milliards d’années, la quantité d’oxygène a très fortement augmenté en peu de temps (environ 10 millions d’années). Cet événement est appelé grande oxygénation de l’atmosphère terrestre. Cette accumulation d’oxygène s’est donc avérée toxique pour certains organismes, qui ont dû s’adapter à ce changement.
Les études des microfossiles montrent les traces des premiers Eucaryotes sur Terre il y a environ 2 milliards d’années. L’hypothèse actuelle expliquant cette apparition est celle de la théorie endosymbiotique. L’histoire aurait débuté lorsqu’une cellule hôte anaérobie, pour laquelle l’oxygène aurait été toxique et ayant la capacité d’assimiler des éléments de son environnement, aurait incorporé une autre cellule aérobie à l’intérieur de son cytoplasme, l’endosymbiote, sans toutefois la digérer. Une relation de bénéfices mutuels se serait mise en place. L’une procurant de l’énergie et une tolérance à l’oxygène en échange d’un habitat protégé et d’un échange mutuel de nutriments. La suite de l’histoire est encore très débattue, il est possible que la cellule hôte ait converti la cellule incorporée en organite ou que des endosymbioses en séries se soient produites, donnant naissances aux mitochondries et aux chloroplastes actuels.
Il existe plusieurs preuves de l’origine procaryotique des mitochondries et des chloroplastes, notamment la présence d’un ADN et de ribosomes propres à ces organites [2]. Pourtant, malgré ces preuves, nous n’avons toujours pas clairement identifié de microorganismes candidats pour être l’hôte originel, ni l’endosymbiote, de cette théorie.
L’hypothèse la plus couramment acceptée aujourd’hui est qu’une archée aurait été l’hôte d’une bactérie, ce qui aurait fait émerger la première cellule eucaryote [3-6]. Depuis longtemps, les chercheurs tentent d’identifier ce premier couple hôte-endosymbiote [*], c’est-à-dire l’ancêtre commun à l’origine des eucaryotes, et de comprendre les mécanismes de sa formation. Aujourd’hui, une étude vient d’identifier une nouvelle archée qui aurait toutes les caractéristiques pour être un hôte possible de la théorie endosymbiotique, ainsi qu’un mécanisme par lequel celle-ci aurait pu incorporer la bactérie symbiotique. Pour la première fois, on pourrait donc avoir une idée un peu plus précise de qui aurait fait quoi, avec qui et comment…
L’hôte : une archée appartenant au groupe d’Asgård
Avant de rentrer dans le vif du sujet, faisons une petite présentation des Archées. Ce domaine du vivant est le moins connu des trois, et pour cause, pendant longtemps les Archées ont été assimilées au domaine des Bactéries, alors appelées Archéobactéries. C’est en 1990 que Carl Woese et ses collaborateurs [7] vont insister sur le fait que les « archéobactéries » sont un groupe fondamentalement différent des bactéries. Ils définissent alors les trois domaines du vivant que l’on connaît actuellement. Le domaine des Archées est créé (Figure 1).
Il existe plusieurs embranchements au sein de ce domaine (dont les noms se terminent presque tous par –archaeota). Celui qui nous intéresse ici est un groupe particulier, décrit récemment, du nom de groupe d’Asgård. La plupart des noms des archées de ce groupe sont basés sur des dieux de la mythologie nordique. Ainsi on trouve : Odin-, Thor-, Loki-, Hel- et Heimdallarchaeota.
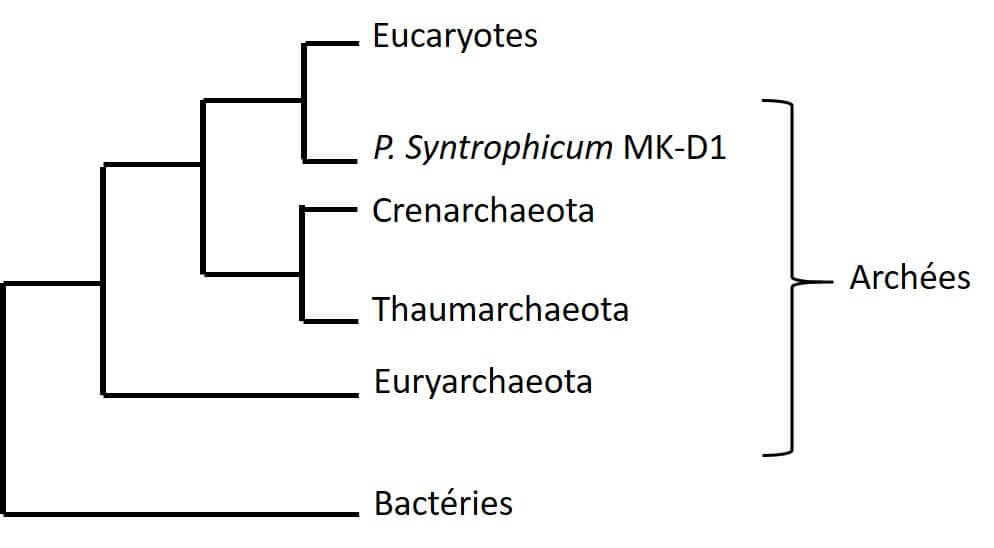
Ce sont les Lokiarchaeota qui ont été découverts les premiers [8] — pour simplifier nous les appellerons Loki — du nom du « Loki’s Castle » (ne cherchez pas un château : il s’agit d’une source hydrothermale entre le Groenland et la Norvège) proche duquel le premier spécimen a été identifié. Ce groupe a, dès sa découverte, suscité l’intérêt des scientifiques en raison de la présence de signatures protéiques que l’on n’avait retrouvées jusqu’alors que chez les Eucaryotes, comme des protéines impliquées dans le trafic membranaire, des vésicules de transport, l’ubiquitine ou encore la formation du cytosquelette. Et encore plus depuis que les auteurs de la publication qui nous intéresse ici ont décrit, en 2020, une nouvelle archée appartenant aux Loki qui aurait les caractéristiques pour être un hôte possible de la théorie endosymbiotique recherchée depuis si longtemps.
L’histoire de la découverte de cette nouvelle archée commence en 2006, lorsqu’une équipe de chercheurs japonais réalise des prélèvements de sédiments sur la crête d’Omine, dans la fosse de Nankai au Japon, à plus de 2 500 m de profondeur. Les premières analyses révèlent assez vite la présence de nombreux microorganismes encore inconnus. En effet, à ce moment-là, les Loki n’avaient pas encore été décrits. L’équipe décide d’essayer de cultiver ces microorganismes inconnus en laboratoire ; ils font alors face à plusieurs défis :
- les archées, en général, sont difficilement cultivables en laboratoire ;
- l’échantillon collecté provient d’une zone anaérobie (= sans dioxygène) et riche en méthane : il faut donc reproduire ces conditions.
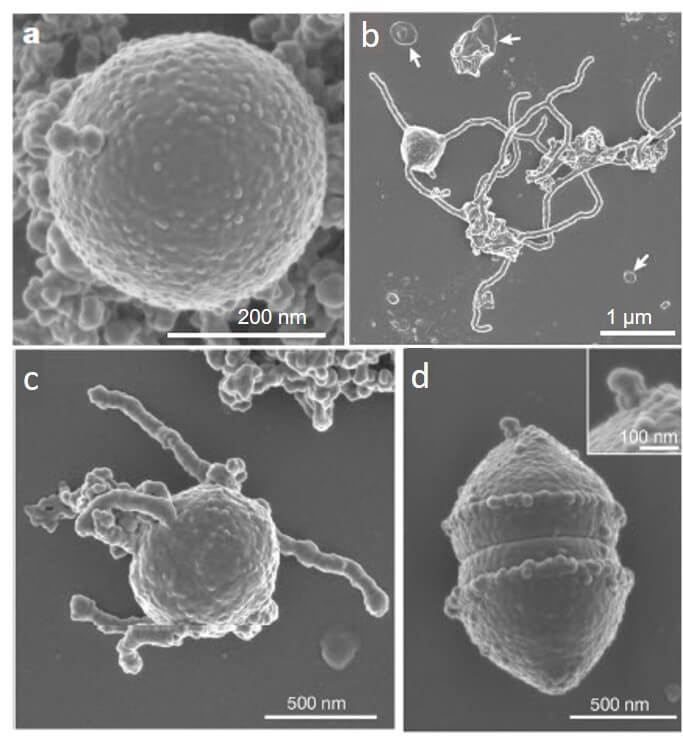
Après plus de 2 000 jours (soit presque 5 ans et demi) de culture dans un bioréacteur enrichi avec les sédiments collectés, l’équipe réussit à obtenir en culture des microorganismes phylogénétiquement divers. Afin de les isoler, l’équipe poursuit les expériences par des cultures et des isolements supplémentaires. Ainsi, dans une culture avec un substrat simple, en milieu anaérobie, supplémenté en protéines de lait et en présence de quatre antibiotiques (les travaux précédents de l’équipe ayant démontré que certaines Archées était tolérantes aux antibiotiques utilisés [9]), l’équipe identifie, après plus de 10 années de recherche, une archée encore inconnue, appelée dans un premier temps MK-D1 (Figure 2).
À partir de là, l’équipe multiplie les expérimentations pour identifier plus précisément les caractéristiques morphologiques, physiologiques et le métabolisme de MK-D1 :
- mise en culture de l’archée sur différents substrats et avec différents partenaires procaryotes afin d’identifier les conditions et le temps de croissance, ainsi que son métabolisme ;
- microscopie électronique à transmission et à balayage pour caractériser la biologie et la physiologie de MK-D1 ;
- analyses ADN pour connaître le génome de MK-D1.Ce dernier point a montré la présence de 80 signatures protéiques eucaryotes, mais que l’on retrouve aussi chez d’autres archées du groupe d’Asgård. De plus, il existe une relation phylogénétique très proche entre MK-D1 et les Eucaryotes, au point que MK-D1 représente le plus proche parent des Eucaryotes découvert à ce jour (Figure 3).
Ainsi, au bout de plus de 10 ans de recherche, l’équipe est parvenue à la caractérisation de cette nouvelle archée, qui s’est révélée appartenir au groupe des archées d’Asgärd et plus précisément aux Lokiarchaeota. Ils lui ont alors donné un nom plus sympa que MK-D1 : Prometheoarchaeum syntrophicum. Ceci en référence au dieu grec Prométhée qui, d’après la mythologie, aurait créé des hommes à partir de boue. Les caractéristiques de P. syntrophicum sont les suivantes :
- petite cellule de forme sphérique, d’environ 550 nm de diamètre, anaérobie et syntrophique ;
- temps de doublement en culture entre 14 et 25 jours ;
- les cellules forment des agrégats ainsi que des vésicules et des protubérances membranaires (« bras » ou « tentacules », Figure 2b et 2c). Ces dernières pourraient leur permettre d’attrapers des bactéries voisines, car il a été observé que ce microorganisme à la particularité de se coller à d’autres bactéries dans les cultures réalisées ;
- présence de protéines eucaryotes dont certaines n’avaient pas encore été observées chez les autres organismes du groupe d’Asgård.
La découverte et la caractérisation d’un nouveau microorganisme est déjà un événement en soi. Mais certaines caractéristiques de P. syntrophicum (protubérances membranaires, protéines eucaryotes et sa capacité à vivre en milieu anaérobie) ont alors suscité la curiosité des scientifiques de l’étude et les ont conduit à pousser leur réflexion plus loin. Ce nouvel organisme réunissait en effet certaines conditions pour que l’on puisse émettre l’hypothèse qu’il pourrait être un hôte possible, encore inconnu, de la théorie endosymbiotique à l’origine de l’émergence des Eucaryotes sur Terre ou au moins un organisme apparenté au LECA : Last Eukaryotic Common Ancestor, le dernier ancêtre commun aux Eucaryotes.
Modèle d’enchevêtrement – engloutissement – endogénéisation pour l’émergence des Eucaryotes (modèle E3)
Les auteurs de la publication ont alors établi un nouveau modèle, baptisé enchevêtrement – engloutissement – endogénéisation (modèle E3), pour lequel il faut deux acteurs, dont voici les caractéristiques :
- un hôte : une archée syntrophique, cellule simple, anaérobie et vivant dans le milieu marin (comme la plupart des organismes à cette époque) ;
- un endosymbiote : une bactérie aérobie (ou au moins tolérante à l’oxygène) et organotrophe.
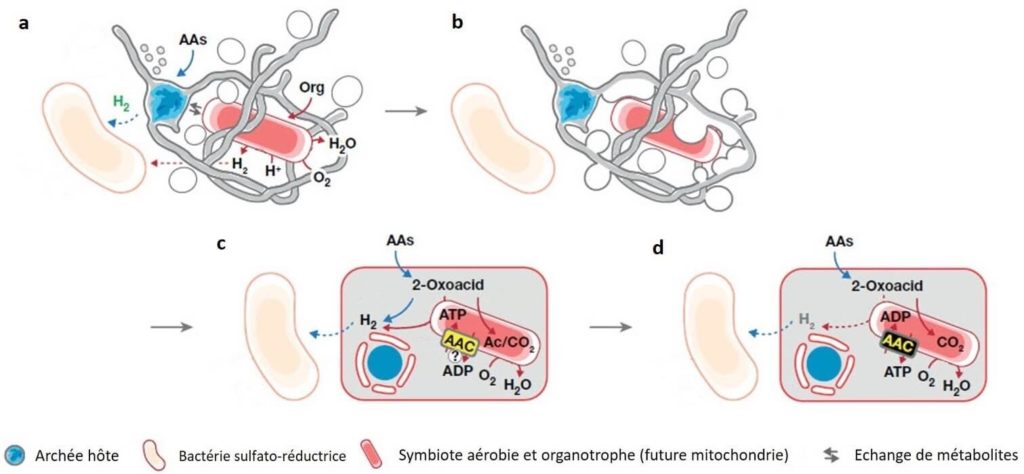
À partir de ce modèle E3, les chercheurs ont proposé une histoire construite et cohérente pouvant expliquer l’origine des Eucaryotes. Selon cette histoire, plusieurs étapes auraient été nécessaires (Figure 4).
- Tout d’abord, une transition entre un mode de vie anaérobie et aérobie. Une relation de dépendance entre deux partenaires s’installe : l’un des acteurs, le futur hôte, dégrade les acides aminés. Les produits secondaires résultants sont utilisés par le partenaire tolérant à l’oxygène, le futur endosymbiote (Figure 4a).
- Ensuite, les structures externes de l’hôte (« bras ou tentacule », Figure 2b et 2c) auraient interagi avec le partenaire symbiotique, améliorant l’interaction physique et « engloutissant » ce dernier. On pourrait dire qu’il l’a « avalé » mais sans le « digérer ». (Figure 4b).
- Suite à cet « engloutissement », l’hôte aurait partagé avec l’endosymbiote des métabolites comme source d’énergie pour ce dernier. En retour, l’endosymbiote consomme l’oxygène environnant et fournit à son tour de l’énergie à l’hôte (sous forme d’ATP) (Figure 4c).
- Enfin, la symbiose entre les deux organismes aurait évolué au cours du temps. Ainsi, à terme, l’hôte aurait délégué le catabolisme et la production d’énergie (ATP) à l’endosymbiote et un canal permettant le passage de ces molécules entre les deux cellules se serait mis en place (canal ADP/ ATP) (Figure 4d). Ce canal, aussi appelé transporteur ADP/ATP, est une protéine majeure de la membrane des mitochondries actuelles.
- Finalement, au cours de l’évolution, des modifications structurelles internes seraient survenues, donnant naissance aux mitochondries actuelles. Il est aussi possible que d’autres endosymbioses se soient produites en série pour parvenir aux Eucaryotes tel que nous les connaissons aujourd’hui.
En conclusion, les auteurs précisent que leur modèle n’est qu’un scénario possible parmi d’autres imaginables pour expliquer l’émergence des Eucaryotes sur Terre. Toutefois, ces travaux apportent de l’eau au moulin à l’hypothèse qu’une archée aurait été l’hôte d’une bactérie, faisant émerger la première cellule Eucaryote. Bien que ce soit une avancée importante, issue de la combinaison de plusieurs types de données (morphologique, métaboliques et génomiques), il est bien sûr nécessaire que les recherches se poursuivent. Tout d’abord sur la connaissance de cette nouvelle Archée mais aussi sur les autres Archées du groupe d’Asgård. Ces microorganismes pourraient fournir de nouvelles informations, notamment comment peut se faire l’engloutissement d’une cellule par une autre, afin de comprendre précisément la chaîne d’évènements qui a conduit à la première cellule Eucaryote.
Vidéo complémentaire (en anglais) :
[*] Il est à noter que l’endosymbiose n’a peut être pas été un épisode unique, ni le fait d’une seule cellule, mais plutôt d’une série d’événements, peut être communs, entre des cellules procaryotes et que les associations bénéfiques se soient maintenues et améliorées par la suite.
[1] Smit M. & Mezger K., Earth’s early O2 cycle suppressed by primitive continents. Nature Geoscience, 2017. DOI : 10.1038/ngeo3030. [Publication scientifique]
[2] Fox, G. E., Origin and evolution of the ribosome. Cold Spring Harb Perspect Biol., 2010. DOI : 10.1101/cshperspect.a003483. [Publication scientifique]
[3] Lopez-Garcia P. & Moreira D., Open questions on the origin of eukaryotes. Trends Ecol. Evol., 2015. DOI : 10.1016/j.tree.2015.09.005. [Publication scientifique]
[4] Martin W. F., et al., Endosymbiotic theories for eukaryote origin. Phil. Trans. R. Soc. Lond., 2015. DOI : 10.1098/rstb.2014.0330. [Publication scientifique]
[5] Koonin E.V., Origin of eukaryotes from within archaea, archaeal eukaryome and bursts of gene gain: eukaryogenesis just made easier? Phil. Trans. R. Soc. Lond. B., 2015. DOI : 10.1098/rstb.2014.0333. [Publication scientifique]
[6] Eme L., et al., Archaea and the origin of eukaryotes. Nature Reviews Microbiology, 2017. DOI : 10.1038/nrmicro.2017.133. [Publication scientifique]
[7] Woese C.R., et al.,Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. PNAS, 1990. DOI : 10.1073/pnas.87.12.4576. [Publication scientifique]
[8] Spang A., et al., Complex archaea that bridge the gap between prokaryotes and eukaryotes. Nature, 2015. DOI : 10.1038/nature14447. [Publication scientifique]
[9] Imachi H., et al., Cultivation of methanogenic community from subseafloor sediments using a continuous-flow bioreactor. ISME Journal, 2011. DOI : 10.1038/ismej.2011.64. [Publication scientifique]
Publié le 21/12/2020
Elodie Billard/Papier-Mâché/CC BY-NC-SA 4.0 2020Texte et images, à l’exception des images soumises à droits d’auteurs différents : voir au cas par cas en légende.